International Journal of Systematic and Evolutionary Microbiology (2009), 59, 2605–2609
Marinobacter szutsaonensis sp. nov., isolated froma solar saltern
Chung-Yi Wang, Chang-Chai Ng, Wen-Sheng Tzeng and Yuan-Tay Shyu
Department of Horticulture, National Taiwan University, 140, Keelung Road, Section 4, Taipei 106,
A Gram-negative, aerobic, non-spore-forming, halophilic bacterial strain, NTU-104T, was isolatedfrom the Szutsao saltern in southern Taiwan, which was previously used as salt production field. The novel isolate grew optimally at 35–40 6C, at pH 7.5–8.0 and in the presence of 5 % (w/v)NaCl. The major fatty acids were C16 : 0, C18 : 1v9c, C16 : 1v9c, C12 : 0 3-OH and C12 : 0. Thepredominant quinone was Q-9. Phosphatidylglycerol, diphosphatidylglycerol andphosphatidylethanolamine were the predominant polar lipids. The DNA G+C content was56.5 mol%. Phylogenetic analyses based on 16S rRNA gene sequences revealed the affiliation ofthe novel isolate to the genus Marinobacter. DNA–DNA hybridization results between strainNTU-104T and the type strains of the most closely related species, Marinobacter pelagius andMarinobacter koreensis, were 36.4 % and 33.2 %, respectively. On the basis of phenotypic,phylogenetic and genetic analyses, strain NTU-104T is considered to represent a novel species ofthe genus Marinobacter. The name Marinobacter szutsaonensis sp. nov. is proposed, with strainNTU-104T (5BCRC 17809T5CGMCC 1.7011T5JCM 15751T) as the type strain.
chemotaxonomic, genetic and phylogenetic characteriza-
Gammaproteobacteria, was first proposed by Gauthier
tion of a halophilic Marinobacter-like strain, NTU-104T,
et al. (1992). The type species of the genus, Marinobacter
hydrocarbonoclasticus, was isolated from sediments col-
A single isolate of strain NTU-104T was obtained from soil
lected in the Gulf of Fos (French Mediterranean coast) at
sediment that was collected from Szutsao saltern, southern
the mouth of a petroleum refinery outlet chronically
Taiwan. The isolate was cultured by the dilution method
polluted by hydrocarbons (Gauthier et al., 1992). Recently,
(Wang et al., 2008). Briefly, soil sediment was dissolved in
many further species, for example Marinobacter gudaonen-
distilled water, diluted 1 : 1 with solutions of 5, 10, 15 and
sis (Gu et al., 2007), Marinobacter salsuginis (Antunes et al.,
20 % (w/v) NaCl and plated on basal medium agar plates.
2007), Marinobacter segnicrescens (Guo et al., 2007),
The pH of the agar plate was adjusted to 7.0 by the
Marinobacter salicampi (Yoon et al., 2007), Marinobacter
addition of 1 M NaOH. The plates were then incubated at
pelagius (Xu et al., 2008), Marinobacter guineae (Montes
37 uC for five days. Single colonies with various morphol-
et al., 2008), Marinobacter psychrophilus (Zhang et al.,
ogies were selected for the growth condition tests. The salt
2008), Marinobacter mobilis and Marinobacter zhejiangensis
requirements of these isolates were determined using basal
(Huo et al., 2008) have been described and, at the time of
medium [l21: 5 g yeast extract (Difco), 5 g Casamino acid
writing, the genus includes 23 recognized species. Species
(Difco), 5 g MgSO4 . 7H2O] with 0–30 % (w/v) NaCl
of this genus are Gram-negative, aerobic, motile, rod-
content. The optimal conditions for growth were deter-
shaped bacteria. All previously described members of this
mined on basal medium with various temperatures (10–
genus have been found to contain C16 : 0v9c, C16 : 1v9c and
70 uC) and pH values (4–10). The growth rate was
C18 : 1 as the predominant fatty acids and to have DNA
monitored using a spectrophotometer at OD660. The
G+C contents ranging from 53 to 59.6 mol% (Liebgott
Gram-reaction was monitored by Gram staining using
et al., 2006). In this study, a morphological, biochemical,
Gram Stain kits (BD), according to the manufacturer’sinstructions. The ability of the novel strain to utilize
Abbreviations: EPS, exopolysaccharides; PHA, poly-b-hydroxyalkanoate.
various carbohydrates and amino acids was tested by usingthe GN2 MicroPlate Identification Test Panel (Biolog). The
The GenBank/EMBL/DDBJ accession number for the 16S rRNA gene
results were read with a MicroPlate reader, using MicroLog
sequence of strain NTU-104T is EU164778.
3.59 software to perform automated reading and iden-
A table detailing the fatty acid contents of strain NTU-104T and some
tification. Tests for the hydrolysis of aesculin, gelatin,
related species of the genus Marinobacter and figures showing the polarlipid analysis and additional phylogenetic trees are available with the
starch, Tween 20 and Tween 80, for oxidase and catalase
reactions and for H2S production were performed as
described by Mata et al. (2002). Enzyme activity was
formed fluorometrically in triplicate using the method of
determined using the API ZYM system (bioMe´rieux).
Sensitivity to antimicrobial agents was determined in basal
Strain NTU-104T was found to be a Gram-negative, motile
medium that contained 50 mg l–1 of each antimicrobial
rod that grew optimally in medium that contained
agent for at least three days. The antimicrobial agents used
approximately 5 % (w/v) salt. This bacterium exhibited
were ampicillin, bacitracin, carbenicillin, cefotaxime,
extreme halotolerance since it was able to grow in a medium
chloramphenicol, erythromycin, kanamycin, nalidixic acid,
containing 0–20 % (w/v) NaCl. M. hydrocarbonoclasticus is
neomycin, nitrofurantoin, novobiocin, nystatin, penicillin,
also an extremely halotolerant species, whereas M. pelagius is
polymyxin B, rifampicin, streptomycin and tetracycline.
a moderately halophilic bacterium with optimal growth at
Cell morphology was observed by scanning electron
5 % NaCl. Strain NTU-104T grew at 10–50 uC and at pH 6–
microscopy. The sample was fixed and processed following
8.5; optimal growth occurred at 35–40 uC and at pH 7.5–
the recommended procedure for preparing specimens
8.0. The new isolate was aerobic and catalase- and oxidase-
(Anto´n et al., 2002). Gold was used to coat the samples
positive. EPS and PHA were not produced. Detailed results
to an approximate thickness of 5 nm. The samples were
are given in the species description.
subsequently observed under a scanning electron micro-scope (Topcon Co.). Exopolysaccharides (EPS) were
The major components of the fatty acids detected in strain
observed by the method of Azeredo & Oliveira (1996).
NTU-104T were C16 : 0 (37.8 %), C18 : 1v9c (23.7 %),
Poly-b-hydroxyalkanoate (PHA) detection was performedusing GC as described by Mas-Castella` & Guerrero (1995).
Table 1. Differential phenotypic characteristics of strain
The G+C content of DNA was determined using the
NTU-104T and other species of the genus Marinobacter
method of Mesbah et al. (1989). The nucleotide mixtureswere separated by HPLC (JASCO) using a Phenomenex
Species: 1, NTU-104T (data from the present study); 2, M. pelagius
C18 column. The set conditions were a flow rate of 1.0 ml
HS225T (data from the present study and Xu et al., 2008); 3, M.
min21 at a temperature of 37 uC, quantified by measure-
koreensis DD-M3T (data from the present study and Kim et al., 2006);
4, M. gudaonensis SL014B61AT (data from the present study and Gu et
al., 2007). All strains are positive in tests for motility, catalase- and
(Sigma) was used as the calibration reference. The
oxidase-activities and for the utilization of acetate and pyruvate. All
composition of the fatty acid methyl esters (FAME) was
strains exhibit cream coloured colonies. All strains are negative for the
analysed using the standard procedure of the Microbial
Identification software (MIDI). The extraction of the fattyacids was performed as described by Heyrman et al. (1999).
Grown cultures were transferred on to trypticase soy agar(TSA) plates, which contained 3 % (w/v) trypticase soy
broth, 1.5 % (w/v) Bacto-Agar (Difco) and 7 % (w/v) NaCl
for 24 h at 37 uC. Single colonies were removed using a
platinum inoculating loop and transferred to 10 ml Teflon
centrifuge tubes with Teflon screw caps (Nalge Nunc
International). FAME profiles were obtained by GLC using
a model 6890N GC (HP) as described by Descheemaeker &
Swings (1995). The identity of the quinones was deter-
mined by HPLC (Shin et al., 1996). Polar lipids were
analysed by two-dimensional TLC, as described by
The nucleic acids of the novel isolates were extracted using
a FastDNA Spin kit (Bio 101), following the manufac-
turer’s instructions. Two universal primers (9F and 1492R)
were used to amplify the 16S rRNA gene (Stackebrandt &
Liesack, 1993). The amplification was performed using a
Amplicons were later sequenced (Mission Biotech) and
aligned with representatives from the genus Marinobacter
and related taxa using multiple sequence alignment
software (CLUSTAL W 1.82; Thompson et al., 1994). A
phylogenetic tree was constructed with the neighbour-
joining, maximum-likelihood and maximum-parsimonyalgorithms with a bootstrap robustness of 1000 using
*Data from: a, Xu et al. (2008); b, Kim et al. (2006); c, Gu et al.
PHYLIP package 3.6b. DNA–DNA hybridization was per-
International Journal of Systematic and Evolutionary Microbiology 59
C16 : 1v9c (10.2 %), C12 : 0 3-OH (9.3 %) and C12 : 0 (7.8 %).
structed by means of the maximum-likelihood and
The fatty acid profile was similar to those of other species
maximum-parsimony algorithms. Although the tree topol-
of the genus Marinobacter, in particular to that of M.
ogies were slightly different, the relationships between the
pelagius (Gu et al., 2007; Guo et al., 2007; Xu et al., 2008)
species of the genus Marinobacter were similar (see
(see Supplementary Table S1, available in IJSEM Online).
Supplementary Figs S3 and S4 in IJSEM Online). Strain
The major isoprenoid quinone of strain NTU-104T was Q9.
NTU-104T exhibited high 16S rRNA gene sequence
Phosphatidylglycerol, diphosphatidylglycerol and phospha-
similarity to M. pelagius JCM 14804T (98.7 %), M. koreensis
tidylethanolamine were the major polar lipids (see
DD-M3T (97.2 %) and M. gudaonensis SL014B61AT
Supplementary Fig. S2 in IJSEM Online). The DNA
(97.0 %). The DNA–DNA relatedness of strain NTU-104T
G+C content of strain NTU-104T was 56.5 mol%, which
to M. pelagius HS225T, M. koreensis DD-M3T and M.
was within the range found previously for recognized
gudaonensis SL014B61AT was 36.4 % (SD53.8 %), 33.2 %
species of the genus Marinobacter (Table 1).
(SD54.1 %) and 30.7 % (SD54.6 %), respectively. All ofthese values were clearly below the 70 % threshold that is
The 16S rRNA gene sequence of strain NTU-104T as
generally used to delineate species. Thus, the 16S rRNA
determined in this study comprised 1435 nt. Comparative
gene sequence data comparisons and DNA–DNA hybrid-
16S rRNA gene sequence analyses showed that strain NTU-
ization results demonstrated that strain NTU-104T was
104T was most closely related to species of the genus
most closely related to the genus Marinobacter but that the
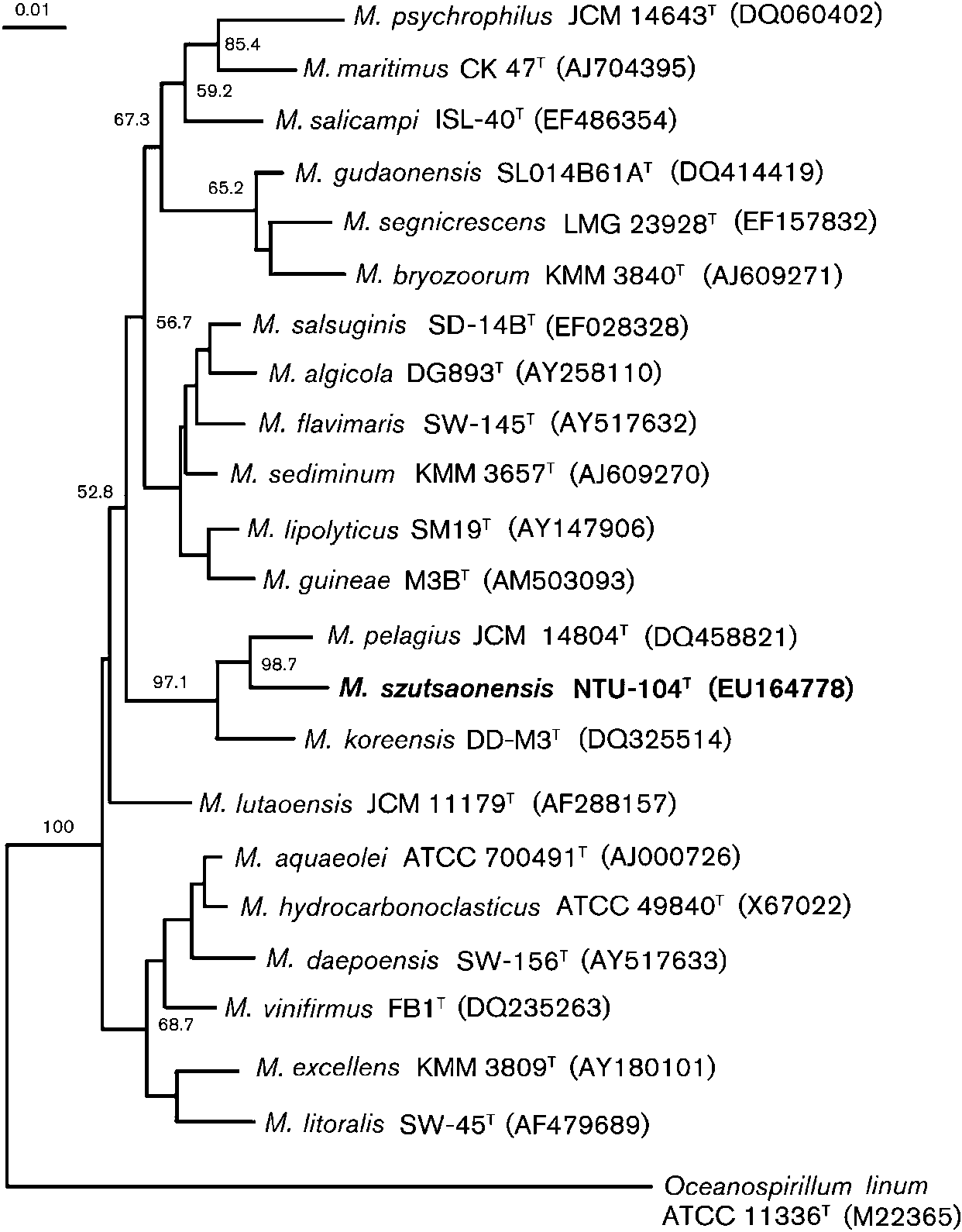
Marinobacter (Fig. 1). Phylogenetic analysis based on 16S
novel strain was not a member of any recognized species of
rRNA gene sequence comparisons showed that strain
this genus. In conclusion, these data, when combined with
NTU-104T formed a coherent cluster with M. pelagius with
the differences revealed by biochemical, physiological and
high bootstrap resampling value (98.7 % by the neighbour-
phylogenetic analyses, are sufficient to suggest that strain
joining method). Other phylogenetic trees were con-
NTU-104T represents a novel species of the genus
Fig. 1. Neighbour-joining tree showing theposition of strain NTU-104T with other speciesof the genus Marinobacter and related taxabased on 16S rRNA gene sequences. Thebootstrap values from 1000 resamplings areindicated at nodes. Bar, 0.01 substitutions persite.
Marinobacter, for which the name Marinobacter szutsao-
Anto´n, J., Oren, A., Benlloch, S., Rodrı´guez-Valera, F., Amann, R. &Rossello´-Mora, R. (2002). Salinibacter ruber gen. nov., sp. nov., a
Description of Marinobacter szutsaonensis
novel, extremely halophilic member of the Bacteria from saltern
crystallizer ponds. Int J Syst Evol Microbiol 52, 485–491.
Marinobacter szutsaonensis (szu.tsao.nen9sis. N.L. masc.
Antunes, A., Franc¸a, L., Rainey, F. A., Huber, R., Nobre, M. F.,
adj. szutsaonensis of szutsao, a former salt field located in
Edwards, K. J. & da Costa, M. S. (2007). Marinobacter salsuginis sp.
nov., isolated from the brine–seawater interface of the Shaban Deep,Red Sea. Int J Syst Evol Microbiol 57, 1035–1040.
Azeredo, J. & Oliveira, R. (1996). A new method for precipitating
2.2 mm) and motile with a single flagellum. Colonies are
bacterial exopolysaccharides. Biotechnol Tech 10, 341–344.
irregular to regular, flat and light translucent and milky
Descheemaeker, P. & Swings, J. (1995). The application of fatty acid
cream colour after 48 h at 37 uC. Growth occurs at NaCl
methyl ester analysis (FAME) for the identification of heterotrophic
concentrations of 0–20 % (w/v), with optimum growth at
bacteria present in decaying Lede-stone of the St. Bavo Cathedral in
5 % NaCl. The optimum growth temperature is 35–40 uC;
Ghent. Sci Total Environ 167, 241–247.
growth is observed at 10–50 uC. The pH range for growth is
Ezaki, T., Hashimoto, Y. & Yabuuchi, E. (1989). Fluorometric
6–8.5, with an optimum at pH 7.5–8.0. Growth occurs
deoxyribonucleic acid-deoxyribonucleic acid hybridization in micro-
under aerobic conditions. Nitrate reduction is positive,
dilution wells as an alternative to membrane filter hybridization in
which radioisotopes are used to determine genetic relatedness among
production is negative. Aesculin, acetate, casein,
bacterial strains. Int J Syst Bacteriol 39, 224–229.
pyruvate, urea and gelatin are hydrolysed, but starch, L-tyrosine, Tween 20, Tween 80, DNA and xanthine are not
Gauthier, M. J., Lafay, B., Christen, R., Fernandez, L., Acquaviva, M.,Bonin, P. & Bertrand, J. C. (1992). Marinobacter hydrocarbonoclasticus
hydrolysed. Oxidase- and catalase-positive. Acid is pro-
gen. nov., sp. nov., a new, extremely halotolerant, hydrocarbon-
duced from D-glucose, D-mannose, maltose, D-mannitol,
degrading marine bacterium. Int J Syst Bacteriol 42, 568–576.
sucrose and D-galactose. Acid is not produced from
Gu, J., Cai, H., Yu, S.-L., Qu, R., Yin, B., Guo, Y.-F., Zhao, J.-Y. & Wu,
cellobiose, D-fructose, melibiose, D-ribose, L-arabinose,
X.-L. (2007). Marinobacter gudaonensis sp. nov., isolated from an oil-
melezitose, D-sorbitol, raffinose, trehalose or D-xylose.
polluted saline soil in a Chinese oilfield. Int J Syst Evol Microbiol 57,
The following substrates are used as sole carbon sources:
L-alanine, fumarate, D-glucose, malate, succinate, D-man-
Guo, B., Gu, J., Ye, Y.-G., Tang, Y.-Q., Kida, K. & Wu, X.-L. (2007).
nose, maltose and sucrose. Melezitose, trehalose, D-salicin,
Marinobacter segnicrescens sp. nov., a moderate halophile isolated
D-sorbitol, D-xylose, benzoate, sorbose, lactose, D-fructose,
from benthic sediment of the South China Sea. Int J Syst Evol
L-tryptophan, formate, L-rhamnose, L-glutamate, glycerol,
gluconate, glutamate, succinate and citrate are not used.
Heyrman, J., Mergaert, J., Denys, R. & Swings, J. (1999). The use of
When assayed with the API ZYM system, alkaline
fatty acid methyl ester analysis (FAME) for the identification of
phosphatase, esterase (C4), leucine arylamidase, valine
heterotrophic bacteria present on three mural paintings showingsevere damage by microorganisms. FEMS Microbiol Lett 181, 55–62.
arylamidase, trypsin, N-acetyl-b-glucosaminidase and a-mannosidase are present, but esterase (C4), esterase lipase
Huo, Y. Y., Wang, C. S., Yang, J. Y., Wu, M. & Xu, X. W. (2008).
Marinobacter mobilis sp. nov. and Marinobacter zhejiangensis sp. nov.,
halophilic bacteria isolated from the East China Sea. Int J Syst Evol
phosphohydrolase, b-galactosidase, a-glucosidase, cystine
arylamidase and a-fucosidase are absent. The type strain is
Kim, B.-Y., Weon, H.-Y., Yoo, S.-H., Kim, J.-S., Kwon, S.-W.,
susceptible to ampicillin, bacitracin, carbenicillin, cefotax-
Stackebrandt, E. & Go, S.-J. (2006). Marinobacter koreensis sp.
ime, chloramphenicol, erythromycin, kanamycin, nalidixic
nov., isolated from sea sand in Korea. Int J Syst Evol Microbiol 56,
acid, nitrofurantoin, nystatin, penicillin, polymyxin B and
tetracycline, but is resistant to neomycin, novobiocin,
Komagata, K. & Suzuki, K. (1987). Lipid and cell-wall analysis in
rifampicin and streptomycin. The predominant isoprenoid
bacterial systematics. Methods Microbiol 19, 161–207.
quinone is Q-9. Phosphatidylglycerol, diphosphatidylgly-
Liebgott, P.-P., Casalot, L., Paillard, S., Lorquin, J. & Labat, M. (2006).
cerol and phosphatidylethanolamine are the predominant
Marinobacter vinifirmus sp. nov., a moderately halophilic bacterium
polar lipids. The major fatty acids are C16 : 0, C18 : 1v9c,
isolated from a wine-barrel-decalcification wastewater. Int J Syst Evol
16 : 1v9c, C12 : 0 3-OH and C12 : 0.
Mas-Castella`, J. & Guerrero, R. (1995). Poly-b-hydroxyalkanoate
The type strain, NTU-104T (5BCRC 17809T5CGMCC
determination in bacteria from aquatic samples. J Microbiol Methods
1.7011T5JCM 15751T), was isolated from Szutsao, a
discarded salt field located in southern Taiwan. The DNA
Mata, J. A., Martinez-Ca´novas, J., Quesada, E. & Be´jar, V. (2002). A
G+C content of the type strain is 56.5 mol%.
detailed phenotypic characterisation of the type strains of Halomonasspecies. Syst Appl Microbiol 25, 360–375.
Mesbah, M., Premachandran, U. & Whitman, W. B. (1989). Precisemeasurement of the G+C content of deoxyribonucleic acid by
This work was supported by Grants from Council of Agriculture and
high-performance liquid chromatography. Int J Syst Bacteriol 39,
International Journal of Systematic and Evolutionary Microbiology 59
Montes, M. J., Bozal, N. & Mercade´, E. (2008). Marinobacter guineae
Wang, C. Y., Chang, C. C., Ng, C. C., Chen, T. W. & Shyu, Y. T. (2008).
sp. nov., a novel moderately halophilic bacterium from an Antarctic
Virgibacillus chiguensis sp. nov., a novel halophilic bacterium, isolated
environment. Int J Syst Evol Microbiol 58, 1346–1349.
from Chigu, a previously commercial saltern located in southern
Shin, Y. K., Lee, J.-S., Chun, C. O., Kim, H.-J. & Park, Y.-H. (1996).
Taiwan. Int J Syst Evol Microbiol 58, 341–345.
Isoprenoid quinone profiles of the Leclercia adecarboxylata KCTC
Xu, X. W., Wu, Y. H., Wang, C. S., Yang, J. Y., Oren, A. & Wu, M. (2008).
1036T. J Microbiol Biotechnol 6, 68–69.
Marinobacter pelagius sp. nov., a moderately halophilic bacterium. Int
Stackebrandt, E. & Liesack, W. (1993). Nucleic acids and classifica-
J Syst Evol Microbiol 58, 637–640.
tion. In Handbook of New Bacterial Systematics, pp. 152–189. Edited
Yoon, J.-H., Lee, M.-H., Kang, S.-H. & Oh, T.-K. (2007). Marinobacter
by M. Goodfellow & A. G. O’Donnell. London: Academic Press.
salicampi sp. now. isolated from a marine solar saltern in Korea. Int J
Thompson, J. D., Higgins, D. G. & Gibson, T. J. (1994).
Syst Evol Microbiol 57, 2102–2105.
improving the sensitivity of progressive multiple sequence alignment
Zhang, D. C., Li, H. R., Xin, Y. H., Chi, Z. M., Zhou, P. J. & Yu, Y. (2008).
through sequence weighting, position-specific gap penalties and
Marinobacter psychrophilus sp. nov., a psychrophilic bacterium
weight matrix choice. Nucleic Acids Res 22, 4673–4680.
isolated from the Arctic. Int J Syst Evol Microbiol 58, 1463–1466.
Ori Better Transcripts Introduction by Dr. Eknoyan GE: Born in 1928 in Haifa, in what was then Palestine, Ori Better witnessed its transfer to what is now Israel. It is there that he graduated from medical school, from the Hadassah Hebrew University medical school, in Jerusalem, in 1957. Inherently bright, and a pioneering spirit, he went on to be on to be one of the founding fathers of nephr
The new Wada anti-doping code comes into effect on 1 January 2004. There are some very significant changes that you need to know about: The major change relates to topical or inhaled corticosteroids. These are banned from Jan 1st 2004, but exemption to use them may be gained via the “therapeutic use simplified a. Topical corticosteroids are widely used for treating many medical conditions.
 C16 : 1v9c (10.2 %), C12 : 0 3-OH (9.3 %) and C12 : 0 (7.8 %).
C16 : 1v9c (10.2 %), C12 : 0 3-OH (9.3 %) and C12 : 0 (7.8 %).