Phylogenetic relationships of african killifishes in the genera aphyosemion and fundulopanchax inferred from mitochondrial dna sequences
Molecular Phylogenetics and EvolutionVol. 11, No. 3, April, pp. 351–360, 1999Article ID mpev.1998.0566, available online at http://www.idealibrary.com on
Phylogenetic Relationships of African Killifishes in the Genera Aphyosemion and Fundulopanchax Inferred from Mitochondrial DNA Sequences Department of Biological Sciences, The University of Tulsa, 600 S. College Avenue, Tulsa, Oklahoma 74104
Received May 12, 1997; revised June 17, 1998
Fundulopanchax, and Nothobranchius) and six mono-
We have analyzed the phylogenetic relationships of
typic genera (Adamus, Foerschichthys, Fundulosoma,52 species representing all defined species groups (J. J. Pronothobranchius, Aphyoplatys, and Episemion). The
Scheel, 1990, Atlas of Killifishes of the Old World, 448
composition and relationships of these genera have under-
pp.) of the African aplocheiloid fish genera Aphyo-
gone numerous changes as our knowledge of these fishes
semion and Fundulopanchax in order to examine their
has grown. The greatest number of changes have occurred
interrelationships and to reveal trends of karyotypic
with regard to the genus Aphyosemion Myers 1924. evolution. The data set comprised 785 total nucleotides
The genus Aphyosemion was originally divided into
from the mitochondrial 12S rRNA and cytochrome b genes. The molecular-based topologies analyzed by
three subgenera: Aphyosemion, Fundulopanchax, and
both maximum parsimony and neighbor-joining sup- Adinops. Those species assigned to Adinops were from
port the monophyly of most previously defined species
east Africa and were later removed to the genus
groups within these two killifish genera. The genus Nothobranchius. The remaining species and subse-
Aphyosemion is monophyletic except for the nested
quent taxa assigned to these subgenera could be di-
position of Fundulopanchax kunzi (batesi group; sub-
vided by distributional criteria and independently by
genus Raddaella) within this clade, suggesting that
phenotypic criteria. The vast majority of these fishes
this taxon was improperly assigned to Fundulopan-
are found in small streams in the understory of the
chax. The remaining Fundulopanchax species sampled
rainforest (Scheel, 1990). The rainforest of equitorial
were supported as being monophyletic in most analy-
Africa is cleanly divided into western and eastern
ses. Relationships among the species groups in both
blocks by the Dahomey Gap, a strip of savanna habitat
genera were not as strongly supported, suggesting that
that extends to the coast in Benin, Togo, and eastern
further data will be required to resolve these relation-
Ghana. In 1966, Clausen recognized the distinctiveness
ships. Additional sampling from the 16S rRNA gene
of those species west of the Dahomey Gap. Subsequent
allowed further resolution of relationships within Fun- dulopanchax, more specifically identifying the nonan-
workers have identified additional morphological char-
nual scheeli group as the basal lineage of this other-
acters that distinguish these western taxa (Zee and
wise annual genus. Chromosomal evolution within
Wildekamp, 1995) and recent DNA sequence data
Aphyosemion has been episodic, with the evolution of a
(Murphy, 1997, Murphy and Collier, 1997, 1999) clearly
reduced n ؍ 9–10 metacentric complement having
identify the western forms as a distinct clade not
occurred in multiple, independent lineages. Polarity of
closely related to the eastern taxa. Thus the remaining
chromosomal reductions within the elegans species
problem with the genus Aphyosemion involves those
group appears to support previous hypotheses concern-
eastern species formerly assigned to the subgenera
ing mechanisms of karyotypic change within the genus Aphyosemion and Fundulopanchax.Aphyosemion.
1999 Academic Press
The subgenus Fundulopanchax was elevated to ge-
neric level by Parenti (1981) based on two characters. Zee and Wildekamp (1995) dispute the diagnostic value
INTRODUCTION
of one of these characters but added four new charac-ters defining Fundulopanchax. Prominent among the
African aplocheiloid killifishes are currently as-
life history traits that distinguish Aphyosemion and
signed to four speciose genera (Aphyosemion, Epiplatys,Fundulopanchax is annualism. Annual fishes (Myers1942, 1955) are those that deposit their eggs in the
substrate where they withstand the dessication of an
Present address: Laboratory of Genomic Diversity, National
Cancer Institute, FCRDC, Frederick, MD 21702.
annual dry season to hatch once the rains resume.
1055-7903/99 $30.00Copyright 1999 by Academic Press
All rights of reproduction in any form reserved.
Members of Fundulopanchax are believed to be annual
while members of Aphyosemion are not (Parenti, 1981;
Taxa Under Study, Proposed Species Groups and Their Abbreviations Used in Figures (Scheel, 1990), and
Chromosome complements are relatively well con-
Various Subgeneric Names Assigned to Specific Taxa
served in teleostean fishes, particularly within theimmense acanthomorph clade (sensu Johnson and Pat-
terson, 1993) in which the predominant diploid karyo-
type is 2n ϭ 48 (Sola et al., 1981). Significant deviations
from this number have occurred in only a handful of
these fish species. One order in particular, the Cypri-
bivittatum,* volcanum, sp. LEC 93/27
nodontiformes (killifishes), displays a striking propen-
sity for clade-specific karyotypic rearrangement. Per-
ahli, australe,* calliurum, celiae
haps the best example is the aplocheiloid genus
Aphyosemion, which shows more inter- and intraspe-
cific chromosomal rearrangements (Scheel, 1990) than
aureum, citrinepinnis, coeleste, occe-
perhaps any other fish genus. Our knowledge of the
types of mechanisms behind karyotypic evolution, and
christyi, cognatum (2), decorsei, elegans
its potential contribution to speciation within this
(3), lamberti, melanopteron, punc-
genus, have been hampered by the lack of a phyloge-
netic framework for this diverse group.
The specific aim of this work was to use mitochon-
drial DNA sequences to assess the monophyly and
composition of Aphyosemion and Fundulopanchax, to
determine the monophyly of recently proposed subgen-
exigoideum, gabunense, louessense,
era and species groups (Scheel, 1990; Table 1) within
these genera, and to determine the polarity of chromo-
somal rearrangements within the molecular phylogeny.
Further, this enlarged data set has allowed further
consideration of the origin of annualism (Murphy and
Collier, 1997) within these genera. We sampled 36
populations of 32 described and 4 undescribed species
of Aphyosemion and 16 species of Fundulopanchax. In
total these represent 14 of Scheel’s (1990) 15 species
groups. The 15th group, composed of a single species,
deltaense, fallax, gulare, schwoiseri,Pronothobranchius kiyawensis, has been subsequently
excluded from Aphyosemion on the basis of both morpho-
logical and molecular characters (Parenti, 1981; Mur-
Note. Generic divisions follow those of Parenti (1981). Numbers in
MATERIALS AND METHODS
parentheses denote number of populations sampled. Data from taxamarked with asterisks have previously been reported (Murphy and
A list of the taxa examined and their sources is in the
Appendix. Mitochondrial DNA was extracted from
muscle or liver tissues. Mitochondrial DNA extractions
and amplification protocols were performed as previ-
ously described (Murphy and Collier, 1996). Some of the
sequences have been previously reported (see Table 1).
We sequenced a 360-bp region of the cytochrome b(cytb) gene and a 425-bp region of the 12S rRNA gene. The primers used were L14724 and H15149 (Kocher et
products were purified with 30,000 MW regenerated
al., 1989; Meyer et al., 1990) for the cytb segment and
cellulose filter devices (Millipore Inc.). Cycle sequenc-
L1091 and H1478 (Kocher et al., 1989) for the 12S
ing using fluorescent-labeled terminators was per-
rRNA segment. Primers 16Sar-L and 16Sbr-H (Palumbi
formed using Ampli-Taq FS DNA polymerase (Applied
et al., 1991) were used to amplify a region of the 16S
Biosystems Inc.). The reactions were purified free of
rRNA gene for Fundulopanchax taxa. The new DNA
fluorescent terminators using Centri-Sep columns
sequences were generated with an automated se-
(Princeton Separations) before loading onto a sequenc-
quencer (ABI 373 Stretch). Symmetric amplification
ing gel (6% Long-Ranger acrylamide, FMC).
PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
Sequences were initially aligned using the program
AF002401. The total analyzed data set consisted of 763
CLUSTAL W (Thompson et al., 1994). Manual adjust-
bp, following removal of 22 bp of unalignable regions
ments were made to the preliminary alignment of the
from the 12S rRNA segment. This resulted in 317
rRNA segments. Regions of length variation due to
variable sites, 265 being parsimony informative. Nucle-
insertions or deletions were omitted from the analyses
otide frequencies for the entire data set were A ϭ 31%,
when they could not be aligned without making assump-
T ϭ 29%, C ϭ 23%, and G ϭ 17% and did not differ
tions concerning homology. The complete aligned data
significantly between taxa. Transition/transversion ra-
sets were analyzed by maximum parsimony (MP) and
tios varied from 1.1 to 19.0 in ingroup comparisons,
neighbor-joining (NJ; Saitou and Nei, 1987) methods.
with many of the higher ratios (particularly among
Parsimony analyses were done with PAUP vers. 3.1.1
closely related taxa) ranging between 5.0 and 10.0.
(Swofford, 1993). In all cases heuristic searches were
Parsimony analysis of the data set when all sites
used (50 replicates, random addition of taxa, TBR
were given equal weight resulted in eight 1806-step
branch-swapping). A series of different weighting
trees having consistency indices of 0.271 and retention
schemes was applied to the parsimony analyses to
indices of 0.565. Figure 1 shows the strict consensus of
adjust for transitional saturation in increasingly diver-
these eight trees. The consistency and retention indices
gent comparisons. This phenomenon is well docu-
are relatively low, most likely attributable to the large
mented in most animal groups, including previous
number of taxa analyzed and the resulting increased
studies of aplocheiloid phylogeny using mitochondrial
probability for homoplasious substitutions at rapidly
DNA (Murphy and Collier, 1996, 1997). We employed
evolving sites. The members of the genus Aphyosemion
the following weighting strategies: (1) all sites given
form a strongly supported group (94% of bootstrap
equal weight; (2) all sites given equal weight, while
replications) which includes Fundulopanchax kunzi
excluding first-codon-position leucine transitions and
nested within this clade. Bootstrap values are also very
all third-codon-position transitions in the cytochrome b
high for nodes defining most of Scheel’s species groups
segment (conservative substitution, CS parsimony; Ir-win et al., 1991); and (3) all transversions weighted five
within Aphyosemion (Fig. 1). The remaining species of
times transitions (5:1 parsimony). These last two
Fundulopanchax form a monophyletic group, though
weighting schemes attempt to resolve deeper diver-
this clade is weakly supported by the bootstrap results
gences due to the greater conservation of the substitu-
(50%). The relationships within Fundulopanchax are
tions analyzed. Bootstrap values for all parsimony
also poorly resolved by these data, with the exception of
analyses were based on 100 heuristic replicates gener-
a few interspecific relationships, which corresponded to
some of Scheel’s (1990) species groups.
Neighbor-joining analyses were generated in MEGA
Weighted parsimony analyses were employed to re-
(vers. 1.01; Kumar et al., 1993) with indels or ambigu-
solve deeper relationships which might be obscured by
ities deleted from the analyses. The Kimura two-
transitional saturation. Both weighting schemes (con-
parameter correction was used to account for transition
servative substitutions and transversions weighted
bias (Kimura, 1980). Confidence probabilities (P
greater than transitions) produced trees in general
Rzhetsky and Nei, 1992) for branches of the NJ tree
agreement with the equal-weighted results (Fig. 1b),
were assessed with the interior-branch test imple-
with most of the species groups within Aphyosemion
mented in MEGA (vers. 1.01, Kumar et al., 1993). We
being monophyletic, though the relationships between
opted to use this method based on recent data suggest-
species groups differed somewhat (see Discussion). The
ing that the bootstrap conservatively underestimates
5:1 parsimony resulted in two trees of 3520 steps (Fig.
the statistical support for groupings within a topology,
1b). This weight was derived from the higher transition
particularly when large numbers of taxa are being
rate among closely related taxa (see above). Results
analyzed (Sitnikova et al., 1995). All trees were rooted
based on weighting transversions between 5 and 10
with members from the genus Nothobranchius (N. kirki
times transitions were equivocal. The analysis based on
and the monotypic subgenus Pronothobranchius kiya-
CS parsimony (eight trees, CI ϭ 0.442, RI ϭ 0.709)
wense) and the allied monotypic genus Fundulosoma
differed primarily from all other analyses in that most
thierryi (Parenti, 1981). A recent molecular analysis of
of the Fundulopanchax taxa (exclusive of F. kunzi) were
the major aplocheiloid genera demonstrated the sister-
not resolved as being monophyletic, collapsing into a
group status of Nothobranchius to a monophyletic
basal polytomy in the consensus tree (not shown). The
Aphyosemion ϩ Fundulopanchax clade (Murphy and
bootstrap trees for these latter two weighted analyses
Collier, 1997), justifying its use here as an outgroup.
gave similar results (shown in Fig. 1b), strongly support-ing most of the species groups within Aphyosemion,
with less resolution between species groups.
The neighbor-joining tree based on Kimura-corrected
Sequences obtained for this study have been depos-
distances (Fig. 2) was congruent with the strongly
ited in GenBank under Accession Nos. AF002284–
supported aspects of the parsimony analyses, with
(a) Consensus tree of eight equal-length cladograms (TL ϭ 1806; CI ϭ 0.271, RI ϭ 0.565) produced when all sites are given equal
weight. Numbers above branches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values below 50% are not shown. Boxesdenote taxa sampled from each of Scheel’s (1990) species groups. Abbreviations for species groups are given in Table 1. (b) Consensus tree oftwo equal-length cladograms (TL ϭ 3519) produced when all transversions were weighted five times greater than transitions. Numbers abovebranches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values in front of the slash are produced by the 5:1transversion/transition weighting scheme. Values following the slash are produced by the bootstrap analysis using conservative substitutions(See text for discussion). Values below 50% are not shown. Boxes denote taxa sampled from each of Scheel’s (1990) species groups. Abbreviations for species groups are given in Table 1.
most of the differences revolving around branches with
To further resolve the relationships with Fundulopan-
low statistical support. Similar to parsimony, the spe-
chax, an additional 472 bp of sequence data was
cies groups within Aphyosemion are strongly supported
obtained from the 16S rRNA gene for each taxon. In
(PC values), while the relationships between the groups
addition, we determined DNA sequences from all three
are weaker. Monophyly of Fundulopanchax, exclusive
gene segments for two Fundulopanchax species not
of F. kunzi, received robust support from the interior-
sampled in the initial portion of this study—F. cinna-
branch test (Fig. 2). Complete deletion of sites with
momeum and F. fallax. Some of the 16S rRNA se-
gaps or the use of different distance corrections changed
quences have been previously reported (Murphy
the topology very little, these differences again being
and Collier, 1997). Trees were rooted with two
observed among branches with low statistical support. Aphyosemion species, and the combined data set (1221
PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
Neighbor-joining tree based on Kimura-corrected distances (Kimura, 1980) with pairwise deletion of gaps. Numbers above the
branches are confidence probabilities based on the interior-branch test implemented in MEGA vers. 1.01 (Kumar et al., 1993). Bars span taxaassigned to Scheel’s (1990) species groups. Dashed lines connect nonadjacent members of these groups. Abbreviations are given in Table 1. Adistance scale is represented at the bottom.
bp, 330 variable sites, 220 of these parsimony informa-
define monophyletic lineages which support Scheel’s
tive) was analyzed by three parsimony methods (all
groupings: bivittatum, calliurum, cameronense, coe-
sites equal weight, conservative substitutions only, and
leste, exiguum, and striatum groups.
transversions weighted five times transitions), neighbor-
The georgiae species group [the subgenus Diapteron
joining, and, as allowed by the smaller size of the data
of Huber and Seegers (1977)] is represented in the
set, maximum likelihood (fastDNAML; Olsen et al.,
combined dataset by a single taxon—A. cyanostictum.
1994). A 6-bp region of the 12S rRNA segment was
Two additional species from this group, abacinum and
deleted due to ambiguity in alignment. Weighted parsi-
georgiae, were sampled; however, the cytochrome b
mony, neighbor-joining, and maximum likelihood analy-
primers were unsuccessful with DNA from these taxa.
ses all produced the topology shown in Fig. 3. Equal-
The results from 12S rRNA gene alone (not shown)
weighted parsimony generated a single tree congruent
demonstrated that all three species comprise a mono-
with Fig. 3, with the exception of F. amieti being placed
phyletic group, with abacinum being basal to the other
basal in the gulare-group clade. The conservative sub-
two. These data also resolved the georgiae group as the
stitution parsimony tree placed the scheeli group sister
sister taxon to the exiguum group, as did the combined
to the gardneri-group/F. ndianum clade.
dataset. The nested placement of this clade within, andnot outside of, Aphyosemion does not support Seeger’s(1980) suggestion of full generic rank for this group. DISCUSSION
Two other Aphyosemion subgenera [Chromaphyo-semion (Radda, 1977) and Kathetys (Huber, 1977)] are
Phylogenetic Relationships within Aphyosemion
also supported here by the apparent monophyly of the
These molecular results are highly concordant with
corresponding bivittatum and exiguum species groups.
previous species-group definitions of the genus Aphyo-
The subgenus Mesoaphyosemion (Radda, 1977), which
semion, created on the basis of karyology, meristics,
includes the cameronense, calliurum, coeleste, elegans,
and geographical distributions (Scheel, 1990). Initial
scheeli, and striatum groups, is clearly not a monophy-
sampling from the following species groups appear to
Phylogenetic hypothesis for the genus Fundulopanchax based on the expanded (cytochrome b ϩ 12S rRNA ϩ 16S rRNA) data set,
using Aphyosemion as an outgroup. Weighted parsimony (TL ϭ 1682), neighbor-joining with Kimura (1980) distances, and maximumlikelihood (Ϫln Likelihood ϭ Ϫ6010.09303) analyses all produced this same topology. The equal-weighted and conservative substitutionparsimony trees are discussed in the text. Numbers above the branches are bootstrap values (500 replicates) compatible with equal-weightedparsimony, conservative substitution parsimony, and 5:1 (Tv:Ts) weighted parsimony. Numbers below the branches are the results of theinterior-branch test (Rzhetsky and Nei, 1992) of the neighbor-joining tree, implemented in MEGA (Kumar et al., 1993).
PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
The placement of the annual Fundulopanchax kunzi
circumpeduncular scale rows characteristic of most
within a larger clade of nonannual Aphyosemion spe-
other Fundulopanchax is also characteristic of the
cies is quite unexpected. This species is part of a small
scheeli group (Zee and Wildekamp, 1995). Given our
group (batesi group) (Scheel, 1990) distributed in up-
topology for Fundulopanchax (Fig. 3) it would appear
land habitats of eastern Cameroon and western Gabon
that 16 or more cp scales is diagnostic for Fundulopan-
disjunct from the usual coastal ranges of the remaining
chax, being secondarily reduced in the derived arnoldi
species of Fundulopanchax. The batesi group is, how-
ever, sympatric in some areas with the groups for whichthe molecular data suggest phylogenetic similarity.
Recently, Zee and Wildekamp (1994) presented new
Basal members of both Aphyosemion (calliurum
morphological characters defining Fundulopanchax
group) and Fundulopanchax (scheeli group) occupy the
which are absent in members of the batesi group
eastern region of Cameroon, suggesting that this region
(subgenus Raddaella). Chorionic puncti, present on the
may have been the center of diversification of these two
surface of eggs of all annual Fundulopanchax species,
genera. The genus Aphyosemion subsequently diversi-
are lacking in the batesi group. All species of Fundulo-
fied to the east with the terminal elegans group rela-
panchax (with the exception of the arnoldi group) have
tively recently filling the Zaire basin. The genus Fundu-
a mean of 16 or more circumpeduncular scales (cp
lopanchax diversified westward in essentially coastal
scales), while all species of Aphyosemion sampled (in-
habitats with the gardneri group expanding inland to
cluding the batesi group) have a mean of 14.2 or fewer
fill much of the area of Nigeria. Fundulopanchax
cp scales. Together, these two morphological characters
walkeri is the only member of the group to have crossed
support the placement of the batesi group within
the Dahomy Gap to occupy western habitat in Ghana
and Togo. Given its rather terminal position within the
The interrelationships between the Aphyosemion spe-
combined Aphyosemion/Fundulopanchax clade, this
cies groups are less apparent with these data. Our
has been a relatively recent event. As such, it does not
results distinguish two major components within the
invalidate the presumption of the epicontinental seas
genus: (1) a strongly supported monophyletic clade
being the viacariant event separating the eastern and
containing the batesi, cameronense, coeleste, and el-
western aplocheiloid taxa (Murphy and Collier, 1997;
egans groups, plus the ungrouped A. labarrei; and (2) a
basal grade composed of the bivittatum, calliurum,exiguum, georgiae, and striatum groups. The former
group is centered primarily around the Congo Riversystem and its neighboring drainages in Gabon while
The suite of traits referred to as annualism includes
the latter group is centered primarily in Cameroon and
behavioral components (bottom spawning), morphologi-
cal components (enlarged dorsal and anal fins of males
The neighbor-joining tree also supports a basal bifur-
and features of chorion structure), and developmental
cation between the calliurum/bivittatum/exiguum/
components (embryonic diapauses). Within the subor-
georgiae groups and the remaining species groups,
der Aplocheiloidei, annualism has been hypothesized to
though the monophyly of the former group is supported
have arisen once and then to have been subsequently
by a very short internode, which receives no support
lost and regained (Murphy and Collier, 1997). The more
from the interior-branch test. However, when members
detailed phylogenetic analysis presented here, with a
of Fundulopanchax are used as an outgroup for the NJ
unique annual species (F. kunzi) nested within an
analysis (data not shown) the calliurum group is found
otherwise nonannual clade and the nonannual scheeli
as the most basal Aphyosemion clade, similar to parsi-
group being the basal group of the otherwise faculta-
tively annual Fundulopanchax, suggests that annual-ism is evolutionarily more plastic than once thought. Phylogenetic Relationships within Fundulopanchax
In contrast to Aphyosemion, only two of Scheel’s
Fundulopanchax species groups are supported by the
Acanthomorph fishes have extremely conserved
current molecular data. One of these, the scheeli group,
karyotypes, with the vast majority of taxa having a
is the lone nonannual taxon within Fundulopanchax,
diploid number of 44–48 chromosomes (Sola et al.,
inhabiting a restricted range in Cameroon and south-
1981). This number is particularly well conserved
eastern Nigeria. The monophyly of this distinctive set
within the speciose Percomorpha. While there are a few
of species is strongly supported (bootstrap ϭ 100% all
scattered examples of significant reductions in number
analyses, PC ϭ 0.99) and the addition of the 16S rRNA
throughout the acanthomorph fishes, the Order Cypri-
segment stabilizes the basal position of the scheeli
nodontiformes exhibits more documented inter- and
group in weighted parsimony, neighbor-joining, and
intraspecific variability than perhaps any other fish
maximum likelihood analyses. The elevated number of
group of equal phylogenetic diversity. The suborder
Cyprinodontoidei has well-conserved chromosome num-bers, with the only significant examples of reductioncoming from the Goodeidae (Turner et al., 1985). Withinthe sister-suborder Aplocheiloidei we see an increasedpropensity for karyotypic reduction, the Neotropicalclade Rivulidae showing a few sporadic cases (n ϭ 10for Pterolebias longipinnis, n ϭ 16–17 in a handful oftaxa) amid a larger trend of n ϭ 22–24 chromosomes(Scheel, 1972, 1990; Garcia et al., 1993; Collier, unpub-lished data).
The African genera show by far the greatest karyologi-
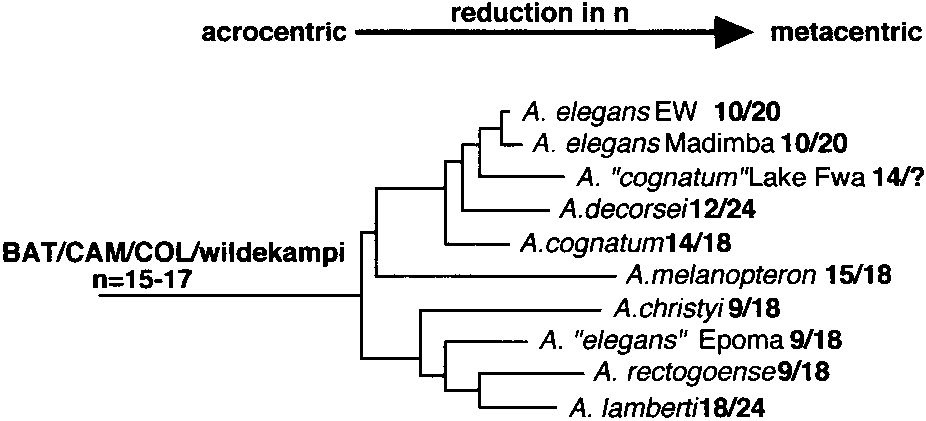
Distribution of karyotypic data onto the phylogeny for the
cal variability—the most striking case found within the
monophyletic Zaire-basin components of the elegans group. Karyo-typic information for each species is from Scheel (1990), except
genus Aphyosemion (Scheel, 1990). Of the eight defined
‘‘elegans’’ Epoma and ‘‘cognatum’’ Lake Fwa (Collier, unpublished
species groups, three (bivittatum, calliurum, and el-egans) contain taxa with karyotypes ranging from n ϭ18 to n ϭ 9 or 10 (Fig. 4). The remaining species groupsshow much less variability in chromosome number and
such reduced chromosome numbers: the bristlemouth
morphology. One of the more notable findings from this
Gonostoma bathyphilum (Gonostomatidae), n ϭ 6; the
study is that the species groups showing reduced
gouramie Sphaerichthys osphromonoides (Belonti-
karyotypes are not phylogenetically restricted. Rather,
idae), n ϭ 8; (Sola et al., 1981).
this propensity toward reduced karyotypes has oc-
Within Aphyosemion a consensus topology can be
curred multiple times in Aphyosemion. A similar ex-
constructed based on the relationships depicted in both
treme reduction in haploid number has occurred in
parsimony and neighbor-joining analyses (Fig. 4). One
Nothobranchius rachovii (n ϭ 9/18 arms; Scheel, 1972,
observation is that a karyotype of 19–20 haploid chro-
1990). This trend is particularly striking because the
mosomes is found in all of the basal species groups
populations having n ϭ 9 to 10 in Aphyosemion (14)
(bivittatum, calliurum, exiguum, georgiae, and stria-
outnumber all other nonaplocheiloid teleosts having
tum), while the monophyletic ‘‘eastern’’ groups (batesi,cameronense, coeleste, and elegans) have an apparentlyancestral upper limit of n ϭ 17 chromosomes. Thepresence of n ϭ 20 chromosomes in the basal groupssuggests that it might be the ancestral karyotype forthe genus and that reduction has occurred in eachgroup. Determining the polarity of chromosome changeswithin most of the species groups is currently notpossible without more complete sampling. However,Scheel’s study of the calliurum group suggests that theNyong-North population of A. ahli (n ϭ 20, acrocentricelements) is ancestral and gave rise to the remainingkaryotypes of reduced number in the group (Scheel,1990). The more derived taxon-pair calliurum andceliae have reduced karyotypes (10 populations, alln Յ 12), and the grouping of these two taxa in allanalyses, together with some analyses showing ahliand australe in basal positions, supports this generaltrend towards reduced karyotypes.
Scheel hypothesized that aplocheiloid karyotypes
have evolved via two major mechanisms: pericentricinversions and centric fusions (Scheel, 1972, 1990). Pericentric inversions would move metacentric centro-meres into terminal positions. Acrocentric chromo-somes could then undergo centric fusions to return thecomplement to one of oversized metacentric elements
Hypothesis for chromosomal evolution within the genus
and a reduced number of chromosomes. If this hypoth-
Aphyosemion. Numbers preceding the slash are haploid chromosome
esis is correct, we would expect to see basal taxa having
numbers; numbers following the slash indicate the number of chromo-
higher chromosome numbers and more acrocentric
some arms in the haploid complement. The range of values for each
elements, while terminal, derived taxa would have
terminal taxon are given on the respective branches. Karyologicaldata are derived from Scheel (1990). See text for discussion.
lower haploid numbers with symmetrical (metacentric)
PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
complements. Our sampling of several described el-ACKNOWLEDGMENTS egans group members presents us with a template fortesting this hypothesis of chromosomal evolution within
We thank the following people for supplying various specimens,
Aphyosemion. Figure 5 shows the NJ topology of the
either wild-caught or aquarium strains: Brian Abbott, Jim Gasior, Liz
elegans group. Analysis of this group alone produces a
Hutchings, Mike Kromrez, Monty Lehman, Robert Langton, Dick
similar topology with both parsimony (all sites equal
Martino, Leonard McCowiac, Craig Rees, Ralph Tepedino, PeteTirbak, Bob Tornatore, Darrell Ullisch, Hans Van Es, and Ed Warner.
weight) and NJ methods. The distribution of karyotype
Many of the aquarium strains were obtained through the American
information onto this tree shows a gradual transition
Killifish Association. We are indebted to this group of naturalists/
from n ϭ 15, mostly acrocentric elements in A. melanop-
hobbyists and various other international killifish organizations for
teron, to both elegans populations having n ϭ 10/18
their past and continuing preservation of these fishes.
arms in the upper lineage. All taxa in the bottomlineage exhibit completely symmetrical complements ofn ϭ 9/18 arms, though there appears to be a rare
REFERENCES
exception of an increase to 18 chromosomes/24 arms inA. lamberti. This single example supports Scheel’s
Agnese, J. F., Romand, R., and Pasteur, N. (1987). Biochemical
differentiation between some African Cyprinodontidae. Z. Zool.
hypothesis; however, more extensive sampling from
Syst. Evol. Forsch. 25: 140–146.
within other groups will be necessary to determine the
Ahl, E. (1924). Zur systematik der altweltlichen Zahnkarpfen der
generality of this pattern of chromosomal evolution.
Unterfamily Fundulidae. Zool. Anz. 40: 49–55.
Brosset, A., and Lachaise, D. (1995). Evolution as a lottery conflicting
APPENDIX
with evolution via sexual selection in African rain forest-dwellingkillifishes (Cyprinodontidae, Rivulinae, Diapteron). In ‘ Evolution-
The following is a list of specimens and their various
ary Biology’’ (M. K. Hecht et al., Eds.), Vol. 28, pp. 217–264.
sources: Aphyosemion ahli, Kribi, Cameroon; A. au-reum, GEB 94/9, Gabon; A. australe, chocolate Aquarium
Clausen, H. S. (1966). Definition of a new cyprinodontid genus and
strain (AS); A. bivittatum, Funge, Cameroon; A. buala-
description of a ‘‘new’’ but well known West African cyprinodont,with clarification of the term ‘ sjoestedi,’’ Aphyosemion sjoestedinum, NDOP, Cameroon; A. calliurum, Epe, Cameroon;
(Lo¨nnberg) and Aphyosemion coeruleum (Boul.). Rev. Zool. Bot.A. cameronense halleri, EMS 90/6 Bikong, Gabon; A.Afric. 74: 331–341. celiae winifrediae, New Butu, Cameroon; A. christyi,
Douchement, J., Romand, R., and Pasteur, N. (1984). Biochemical
HZ 85/8, Zaire; A. citrinepinnis, GEB 94/1, Gabon; A.
differentiation in West African Cyprinodontid fish of the genus
coeleste, RPC/5, Congo Republic; A. cognatum, Band-
Aphyosemion. Biochem. Syst. Ecol. 12: 325–333.
undu, Zaire; A. ‘ cognatum,’’ Lake Fwa, Zaire; A. cyanost-
Felsenstein, J. (1985). Confidence limits on phylogenies: An approach
ictum, Makouko, Gabon; A. decorsei, RCA 91/3, Central
using the bootstrap. Evolution 39: 783–791.
African Republic; A. elegans, ‘‘EW,’’ Naoimda, Zaire; A.
Garcia, G., Scvortzoff, E., Ma´spoli, M. C., and Vaz-Ferreira, R. (1993). elegans, Epoma, Congo Republic; A. elegans, Madimba,
Analysis of karyotypic evolution in natural populations of Cynole-
Zaire; A. exigoideum, N’goudoufola, Gabon; A. exiguum,bias (Pisces: Cyprinodontiformes, Rivulidae) using banding tech-
GKCAR 90/4, Central African Republic; A. gabunense
niques. Cytologia 58: 85–94. boehmi, AS, Gabon; A. labarrei, AS, Zaire; A. lamberti,
Huber, J. H. (1977). Liste nominal annote´e de Aphyosemion Myers,
NRSC, Gabon; A. louessense, Sibiti, Congo Republic; A.
avec description de Raddaella et Kathetys deux sous-genres a` la biologie originale. Suppl Killi Rev. France 4: 1–14. maculatum, LEC93/4, Gabon; A. melanopteron (ϭcongi-
Huber, J. H. (1979). Cyprinodontines de la cuvette congolaise (Ada-cum), AS, Zaire; A. mimbon, LEC93/19, Gabon; A.mus formosus n. gen. n. sp. et nouvelle description de Aphyosemionoccellatum, G-20, Gabon; A. ogoense, RPC/206, Gabon;
splendidum). Rev. Franc¸aise Aquar. 6: 5–10. A. primigenium, 88/6, Gabon; A. punctatum, LEC,
Irwin, D. M., Kocher, T. D., and Wilson, A. C. (1991). Evolution of the
Gabon; A. rectogoense, GAB 90/ABB, Gabon; A. stria-
cytochrome b gene in mammals. J. Mol. Evol. 32: 128–144. tum, Cape Esterias, Gabon; A. volcanum, Kumba,
Johnson, G. D., and Patterson, C. (1993). Percomorph phylogeny: A
Cameroon; A. wildekampi, AS; A. sp. LEC93/27, Gabon;
survey of acanthomorphs and a new proposal. Bull. Mar. Sci. 52: Fundulopanchax amieti, AS, Cameroon; F. cinna-momeum, AS, Cameroon; F. deltaense, AS, Nigeria; F.
Kimura, M. (1980). A simple method for estimating evolutionary
fallax, Mamou, Ghana; F. filamentosum, AS, Nigeria; F.
rates of substitution through comparative studies of nucleotide
gardneri, Akure, Nigeria; F. gulare, AS, Nigeria; F.
sequences. J. Mol. Evol. 29: 170–179. kunzi, CGE/91, Gabon; F. mirabile moense Takwai,
Kocher, T. D., Thomas, W. K., Meyer, A., Edwards, S. V., Pa¨a¨bo, S.,
Cameroon; F. ndianum, AS, Nigeria; F. robertsoni, AS,
Villablanca, F. X., and Wilson, A. C. (1989). Dynamics of mitochon-
Cameroon; F. oeseri (ϭsantaisabellae), AS, Bioko Is-
drial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 86: 6196–6200.
land; F. scheeli, AS, Nigeria; F. schwoiseri, AS, Cam-
Kottelat, N. (1976). Modifications taxonomiques au sein des super-
eroon; F. sjoestedi, AS, Nigeria; F. walkeri, AS; Fundulo-
espe`ces Aphyosemion gardneri (Blgr. 1911) et A. walkeri (Boul. soma thierryi, AS; Nothobranchius kirki, Chilwa,
1911) avec une espe`ce et une sous-espe`ce ‘‘nouvelle’’ mais connues
Malawi; Pronothobranchius kiyawense, AS. Collection
et une sous-genre nouveau. Aphyosemion (Fundulopanchax) walk-
codes are detailed in Langton (1996). eri (Boulenger, 1911). Aquarama 22: 157–166.
Kumar, S., Tamura, K., and Nei, M. (1993). MEGA: Molecular
¨ rzl, E. (1987). ‘‘Colour Atlas of the Cyprinodonts
Evolutionary Genetics Analysis, vers. 1.01. Institute of Molecular
of the Rain Forest of Tropical Africa,’’ Verlag Otto Hoffmann,
Evolutionary Genetics, Pennsylvania State University.
Langton, R. W. (1996). ‘‘Wild collections of Killifish, 1950–1995.’’
Rzhetsky, A., and Nei, M. (1992). A simple method for estimating and
American Killifish Association, 70 pp.
testing minimum-evolution trees. Mol. Biol. Evol. 9: 945–967.
Meyer, A., Kocher, T. D., Basasibwaki, P., and Wilson, A. C. (1990).
Saitou, N., and Nei, M. (1987). The neighbor-joining method: A new
Monophyletic origin of Lake Victorian cichlid fishes suggested by
method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4:
mitochondrial DNA sequences. Nature 347: 550–553.
Murphy, W. J. (1997). Molecular Systematics and Biogeography of
Scheel, J. J. (1968). ‘‘Rivulines of the Old World,’’ T. F. H. Publ., Inc.,
Fishes of the Suborder Aplocheiloidei (Atherinomorpha, Cypri-
nodontiformes). Ph.D. dissertation, The University of Tulsa. Univer-
Scheel, J. J. (1972). Rivuline karyotypes and their evolution (Rivuli-
sity Microfilms, Ann Arbor, Michigan. 239 pp.
nae, Cyprinodontidae, Pisces). Z. Zool. Syst. Evol. Forsch. 10:
Murphy, W. J., and Collier, G. E. (1996). Phylogenetic relationships
within the aplocheiloid fish genus Rivulus (Cyprinodontiformes,
Scheel, J. J. (1990). ‘‘Atlas of Killifishes of the Old World,’’ T. F. H.
Rivulidae): Implications for Caribbean and Central American
biogeography. Mol. Biol. Evol. 13: 642–649.
Sitnikova, T., Rzhetsky, A., and Nei, M. (1995). Interior-branch and
Murphy, W. J., and Collier, G. E. (in press). A molecular phylogeny for
bootstrap tests of phylogenetic trees. Mol. Biol. Evol. 12: 319–333.
aplocheiloid fishes (Atherinomorpha, Cyprinodontiformes): The
Sola, L., Cataudella, S., and Capana, E. (1981). New developments in
role of vicariance and the origins of annualism. Mol. Biol. Evol. 14:
vertebrate cytotaxonomy III. Karyology of bony fishes: A review. Genetica 54: 285–328.
Murphy, W. J., Nguyen, T. N. P., Taylor, E. B., and Collier, G. E.
Swofford, D. L. (1993). PAUP-Phylogenetic analysis using parsimony,
(1999). Mitochondrial DNA phylogeny of West African aplocheiloid
vers. 3.1.1. Illinois Natural History Survey, Champaign.
killifishes (Cyprinodontiformes, Aplocheilidae). Mol. Phy. Evol. 11: 343–350.
Tamura, K. (1992). Estimation of the number of nucleotide substitu-
tions when there are strong transition–transversion and G ϩ C
Myers, G. S. (1924). A new poeciliid from the Congo with remarks on
content biases. Mol. Biol. Evol. 9: 678–687.
funduline genera. Am. Mus. Novit. 116: 1–9.
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL
Myers, G. S. (1933). The classification of the African cyprinodont
W: Improving the sensitivity of progressive multiple sequence
fishes with a discussion of the geographical distribution of the
alignment through sequence weighting, position specific gap penal-
Cyprinodontidae of the world. Stanford Univ. Bull. 8: 10–12.
ties and weight matrix choice. Nucleic Acids Res. 22: 4673–4680.
Myers, G. S. (1942). Studies on South American freshwater fishes I.
Turner, B. J., Grudizen, T. A., Adkisson, K. P., and Worrell, R. A. Stanford Ichthyol. Bull. 2: 89–124.
(1985). Extensive chromosomal divergence within a single river
Myers, G. S. (1952). Annual fishes. Aquarium J. (San Francisco) 23:
basin in the Goodeid Fish, Ilyodon furcidens. Evolution 39:
Parenti, L. R. (1981). A phylogenetic and biogeographic analysis of
Wildekamp, R. H. (1993). ‘ A World of Killies: Atlas of the Oviparous
cyprinodontiform fishes (Teleostei; Atherinomorpha). Bull. Am.
Cyprinodontiform Fishes of the World,’’ Vols. I and III, American
Mus. Nat. Hist. 168: 335–557.
Killifish Association, Inc., Mishawaka, IN.
Radda, A. C. (1969). Fundulosoma thierryi und ihre Verwandten.
Zee, J. R. van der, and Wildekamp, R. H. (1995). Description of a new
Aquaria 16: 1–5. Fundulopanchax species (Cyprinodontiformes: Aplocheilidae) from
Radda, A. C. (1977). Vorlaufige Beschreibung von vier neuen Subgen-
the Niger delta, with a redefinition of the genus Fundulopanchax.
era der Gattung Aphyosemion Myers. Aquaria 24: 209–216. J. African Zool. 108: 417–434.
Oyster Creek Talking Points I urge the NJDEP to protect the Barnegat Bay by requiring Exelon Corporation, owner of Oyster Creek Nuclear Power Plant to use best available technology to control the plant’s environmental impacts by installing a closed loop cooling tower. This technology is a cost-effective way to stop extensive environmental damage caused by the power plant’s current “once-thro
Inspirational Times It’s About You! K a i s e r P e r m a n e n t e E m p l o y e e W e l l n e s s P r o g r a m MAY IS AMERICAN STROKE MONTH DO YOU KNOW THE SIGNS AND SYMPTOMS OF STROKE? What is a stroke? Brain damage can begin within Symptoms of a stroke hap-the damaged area of the brain What are the symptoms? WHAT CAN YOU DO TO CONTROL YOUR ALLERGY SYMPTOMS?

 Molecular Phylogenetics and EvolutionVol. 11, No. 3, April, pp. 351–360, 1999Article ID mpev.1998.0566, available online at http://www.idealibrary.com on
Phylogenetic Relationships of African Killifishes in the
Molecular Phylogenetics and EvolutionVol. 11, No. 3, April, pp. 351–360, 1999Article ID mpev.1998.0566, available online at http://www.idealibrary.com on
Phylogenetic Relationships of African Killifishes in the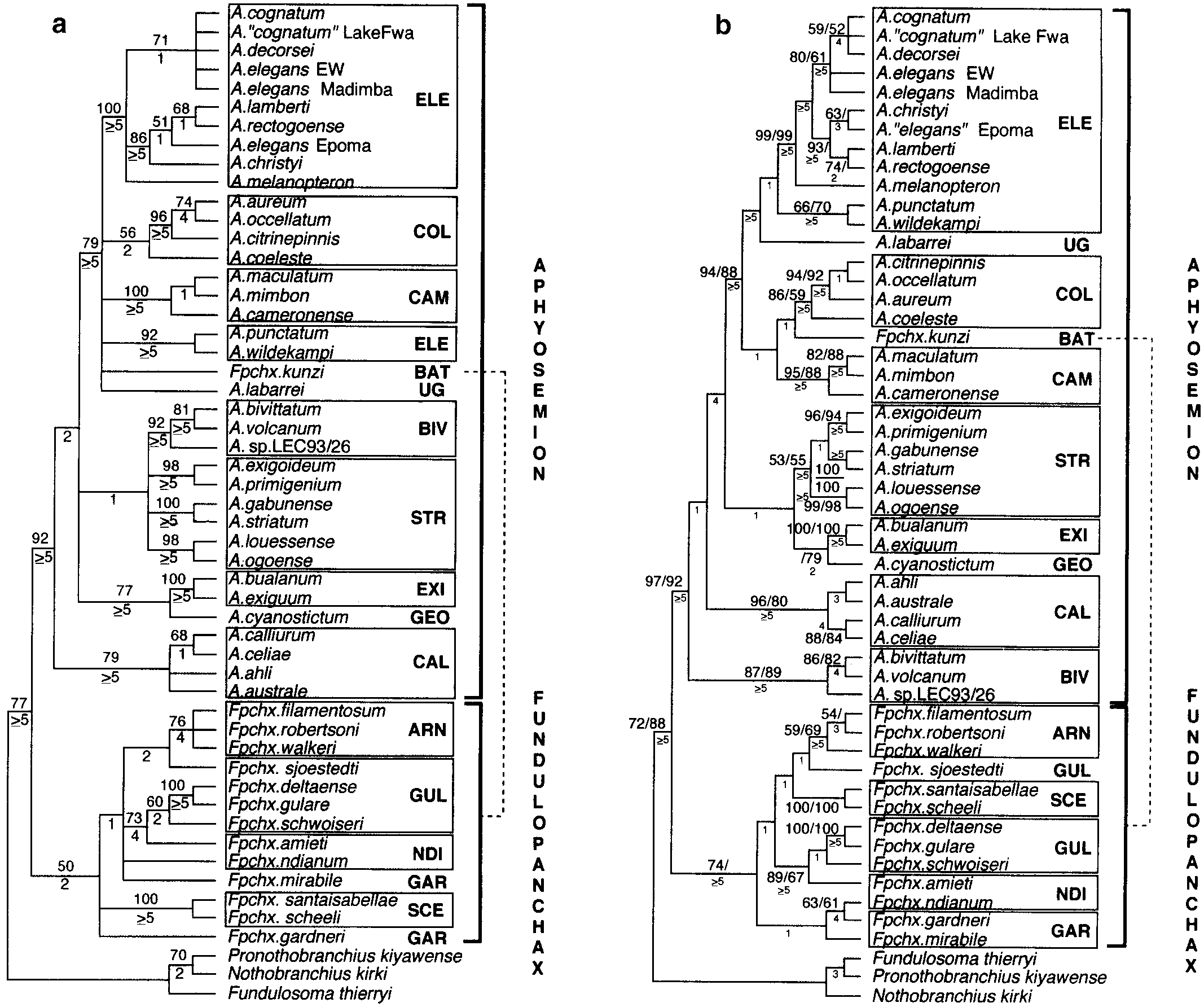 (a) Consensus tree of eight equal-length cladograms (TL ϭ 1806; CI ϭ 0.271, RI ϭ 0.565) produced when all sites are given equal
weight. Numbers above branches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values below 50% are not shown. Boxesdenote taxa sampled from each of Scheel’s (1990) species groups. Abbreviations for species groups are given in Table 1. (b) Consensus tree oftwo equal-length cladograms (TL ϭ 3519) produced when all transversions were weighted five times greater than transitions. Numbers abovebranches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values in front of the slash are produced by the 5:1transversion/transition weighting scheme. Values following the slash are produced by the bootstrap analysis using conservative substitutions(See text for discussion). Values below 50% are not shown. Boxes denote taxa sampled from each of Scheel’s (1990) species groups.
(a) Consensus tree of eight equal-length cladograms (TL ϭ 1806; CI ϭ 0.271, RI ϭ 0.565) produced when all sites are given equal
weight. Numbers above branches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values below 50% are not shown. Boxesdenote taxa sampled from each of Scheel’s (1990) species groups. Abbreviations for species groups are given in Table 1. (b) Consensus tree oftwo equal-length cladograms (TL ϭ 3519) produced when all transversions were weighted five times greater than transitions. Numbers abovebranches are bootstrap values (Felsenstein, 1985) based on 100 replications. Values in front of the slash are produced by the 5:1transversion/transition weighting scheme. Values following the slash are produced by the bootstrap analysis using conservative substitutions(See text for discussion). Values below 50% are not shown. Boxes denote taxa sampled from each of Scheel’s (1990) species groups.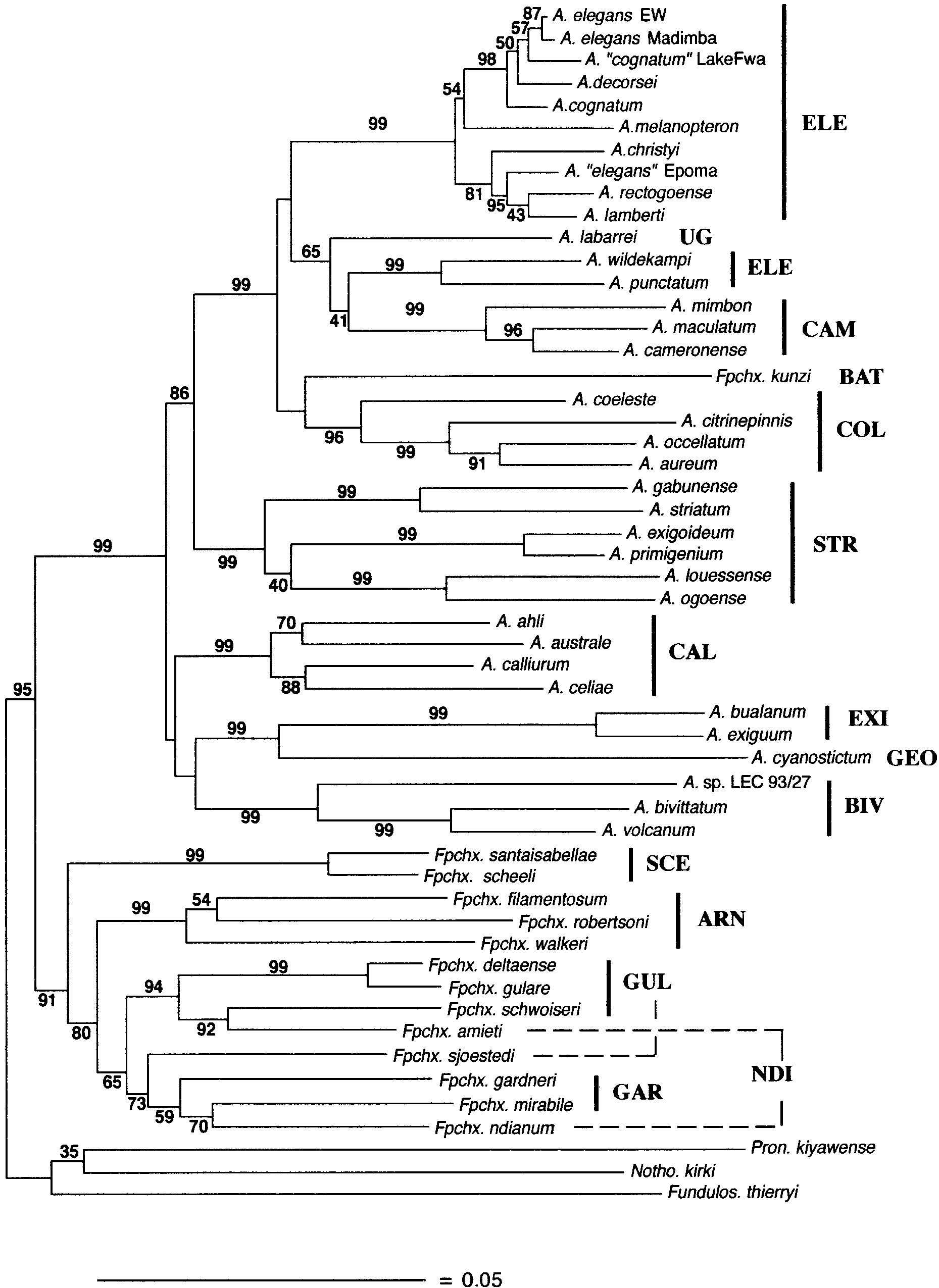 PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
Neighbor-joining tree based on Kimura-corrected distances (Kimura, 1980) with pairwise deletion of gaps. Numbers above the
branches are confidence probabilities based on the interior-branch test implemented in MEGA vers. 1.01 (Kumar et al., 1993). Bars span taxaassigned to Scheel’s (1990) species groups. Dashed lines connect nonadjacent members of these groups. Abbreviations are given in Table 1. Adistance scale is represented at the bottom.
PHYLOGENETIC RELATIONSHIPS OF AFRICAN KILLIFISHES
Neighbor-joining tree based on Kimura-corrected distances (Kimura, 1980) with pairwise deletion of gaps. Numbers above the
branches are confidence probabilities based on the interior-branch test implemented in MEGA vers. 1.01 (Kumar et al., 1993). Bars span taxaassigned to Scheel’s (1990) species groups. Dashed lines connect nonadjacent members of these groups. Abbreviations are given in Table 1. Adistance scale is represented at the bottom.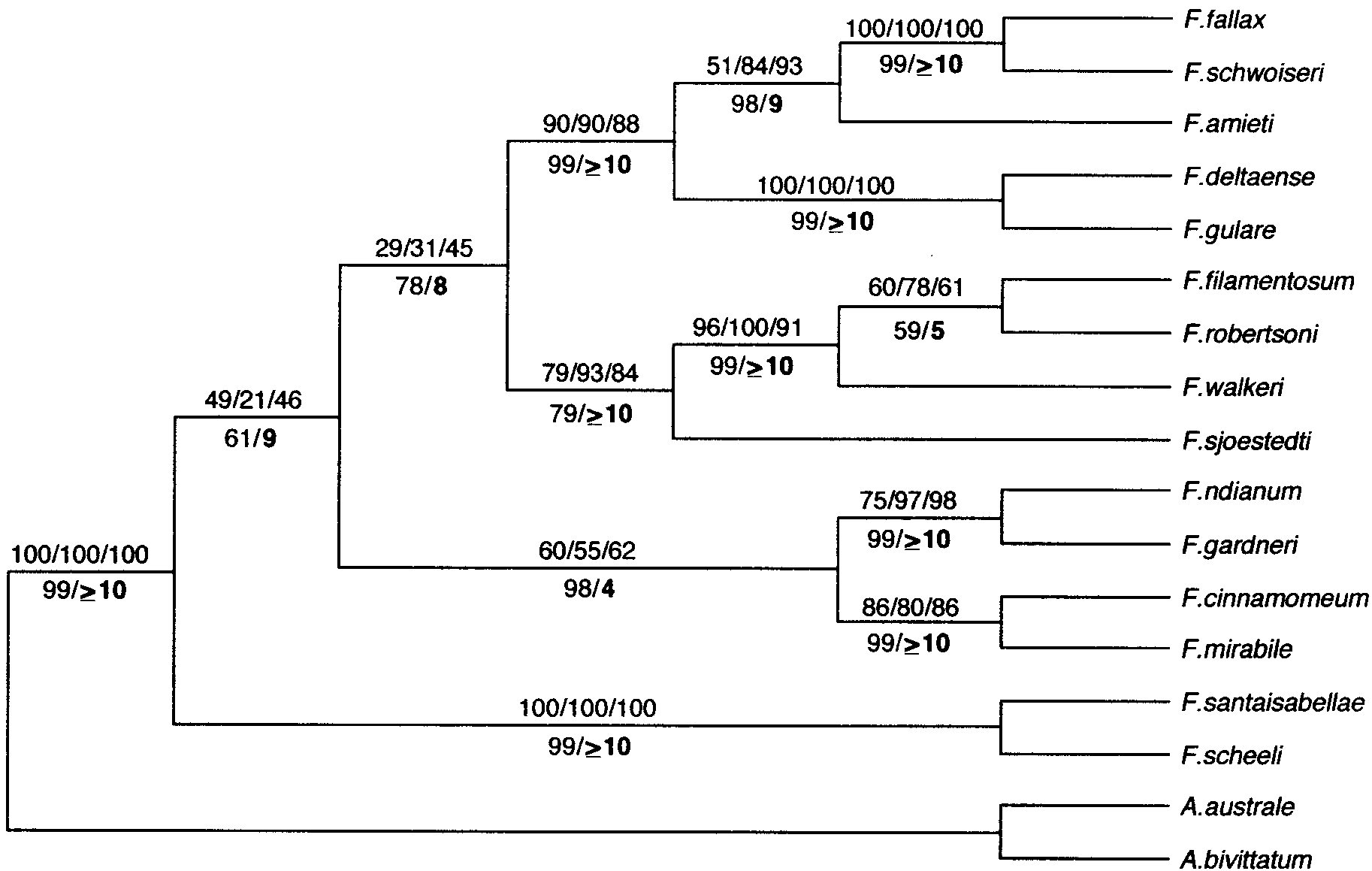 bp, 330 variable sites, 220 of these parsimony informa-
define monophyletic lineages which support Scheel’s
tive) was analyzed by three parsimony methods (all
groupings: bivittatum, calliurum, cameronense, coe-
sites equal weight, conservative substitutions only, and
leste, exiguum, and striatum groups.
bp, 330 variable sites, 220 of these parsimony informa-
define monophyletic lineages which support Scheel’s
tive) was analyzed by three parsimony methods (all
groupings: bivittatum, calliurum, cameronense, coe-
sites equal weight, conservative substitutions only, and
leste, exiguum, and striatum groups.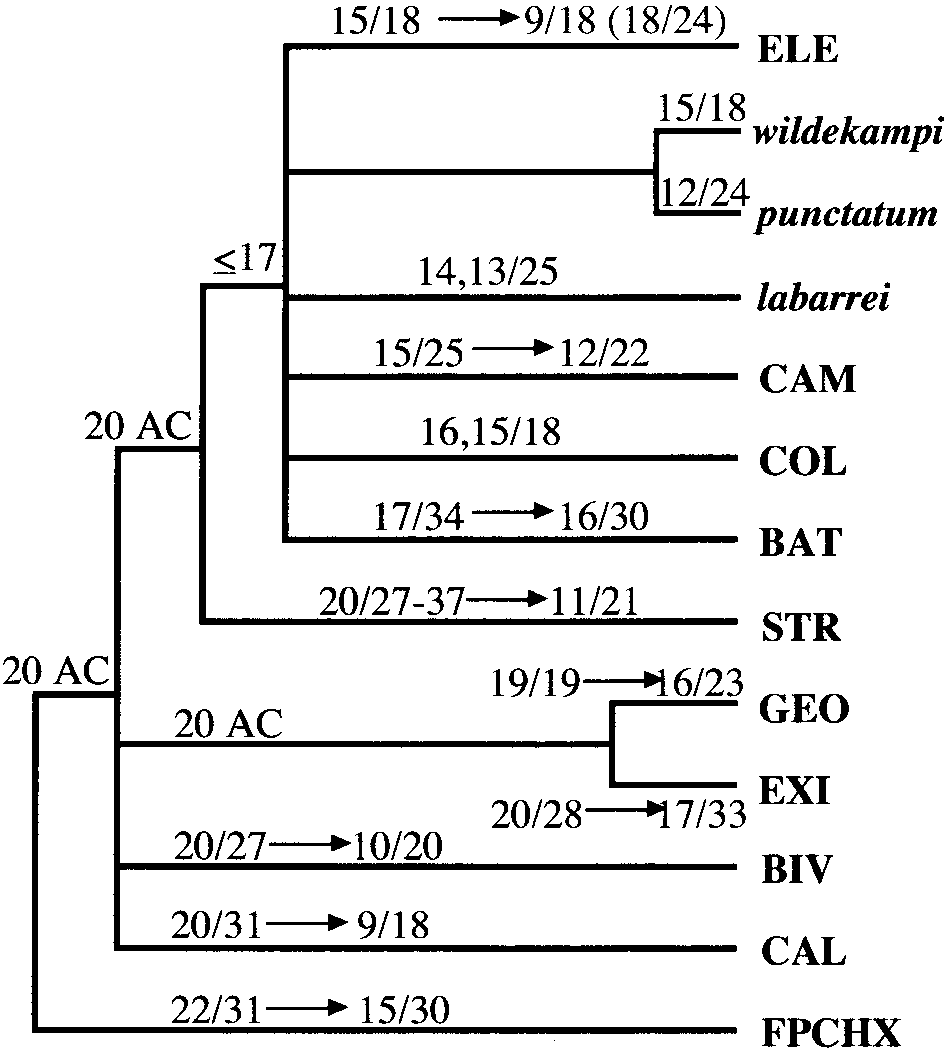
 Cyprinodontoidei has well-conserved chromosome num-bers, with the only significant examples of reductioncoming from the Goodeidae (Turner et al., 1985). Withinthe sister-suborder Aplocheiloidei we see an increasedpropensity for karyotypic reduction, the Neotropicalclade Rivulidae showing a few sporadic cases (n ϭ 10for Pterolebias longipinnis, n ϭ 16–17 in a handful oftaxa) amid a larger trend of n ϭ 22–24 chromosomes(Scheel, 1972, 1990; Garcia et al., 1993; Collier, unpub-lished data).
Cyprinodontoidei has well-conserved chromosome num-bers, with the only significant examples of reductioncoming from the Goodeidae (Turner et al., 1985). Withinthe sister-suborder Aplocheiloidei we see an increasedpropensity for karyotypic reduction, the Neotropicalclade Rivulidae showing a few sporadic cases (n ϭ 10for Pterolebias longipinnis, n ϭ 16–17 in a handful oftaxa) amid a larger trend of n ϭ 22–24 chromosomes(Scheel, 1972, 1990; Garcia et al., 1993; Collier, unpub-lished data).