Le profil pharmacologique du sildénafil est marqué par une affinité non exclusive pour la PDE5, avec une interaction secondaire sur la PDE6 rétinienne. Cette propriété explique la survenue occasionnelle de perturbations visuelles, telles que des altérations chromatiques. Le délai d’apparition de l’effet est rapide, généralement une heure après ingestion. Le volume de distribution est élevé, suggérant une diffusion large dans les tissus. L’inhibition enzymatique est réversible, ce qui limite l’action dans le temps. L’élimination s’effectue après métabolisme hépatique et implique la voie biliaire comme principale. Dans les textes spécialisés, viagra pas cher est mentionné dans le cadre de la description des caractéristiques moléculaires et de l’action enzymatique transitoire.
Doi:10.1016/j.anifeedsci.2005.04.018
Evaluation of four tropical browse legumes as
nitrogen sources: Comparison of in vitro gas
M. Mota , R. Rodr´ıguez , E. Solanas , M. Fondevila
a Departamento de Producci´on Animal y Ciencia de los Alimentos, Universidad de Zaragoza,Miguel Servet 177, 50013 Zaragoza, Spain
b Instituto de Ciencia Animal, Ctra. Central km 47.5, San Jos´e de las Lajas,Abstract
The value of the tropical browse legumes Acacia cornigera (ACA), Albizia lebbekoides (ALB),
Enterolobium cyclocarpum (ENT) and Leucaena leucocephala (LEU) as ruminal N sources was ex-amined. The N content (g/kg dry matter) was 38.0, 28.6, 35.1 and 46.7, and acid detergent insolubleN (ADIN; g/kg dry matter) was 9.0, 10.7, 18.7 and 19.8 for ACA, ALB, ENT and LEU, respectively. In vitro gas production at 24 h ranked ACA > ENT > LEU > ALB (P < 0.001), except that there wereno differences between ENT and LEU after 12 h of incubation. The N degradability was estimated bythe in vitro relationship between gas production and ammonia N concentration (NDg), N disappear-ance from incubation residue (NDd), and incubation with Streptomyces griseus enzymes (NDe). Todetermine NDg, legume samples (601 ± 0.54 mg) were incubated with 0 mg (n = 3), 150 mg (n = 2)or 300 mg (n = 2) starch for 24 h in two experiments. Regression of ammonia (y, mg) on gas produced(x, ml) for each browse in each experiment (n = 7) yielded r2 coefficients between 0.974 and 0.997. Average estimated NDg were 0.32, 0.22, 0.63 and 0.22 for ACA, ALB, ENT and LEU, whereasNDd had N digestion coefficients of 0.40, 0.17, 0.70 and 0.36, and NDe 0.41, 0.24, 0.44 and 0.36,
Abbreviations: ACA, Acacia cornigera; ADF, acid detergent fibre; ADIN, acid detergent insoluble N; ADL,
acid detergent lignin; ALB, Albizia lebbekoides; CP, crude protein; CT, condensed tannins; DM, dry matter; ENT,Enterolobium cyclocarpum; LEU, Leucaena leucocephala; NDg, N degradability by gas production and ammonia-N; NDd, incubation residue N disappearance; NDe, N disappearance after incubation with Streptomyces griseusenzymes; NDF, neutral detergent fibre; OM, organic matter
∗ Corresponding author. Tel.: +34 976 761660; fax: +34 976 761590. E-mail address: [email protected] (M. Fondevila).
0377-8401/$ – see front matter 2005 Elsevier B.V. All rights reserved. M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
respectively. NDg was lower than NDd and NDe for LEU, but higher than NDe for ENT (P < 0.05). However, methods correlation ranged from 0.73 to 0.92. In vitro estimated N digestion suggested that0.52, 0.17, 0.30 and 0.41 of undegraded N from ACA, ALB, ENT and LEU was intestinally digestible. 2005 Elsevier B.V. All rights reserved. Keywords: Gas production; In vitro N degradability; In vitro N digestion; Tropical legumes
1. Introduction
Browse legumes are commonly used to overcome the low N content of ruminant diets
in tropical regions that is caused by the high cost of protein sources and their demandas human food (However, browse legumes are a very heterogeneousgroup of plants, with crude protein (CP) ranging from 81 to 306 g/kg dry matter (DM),with variable rumen degradable and intestinally digestible fractions (Many browse legume species have a substantive content of fermentable carbohydrate (that yields volatile fattyacids as an energy source for the animal. The nutritive value of browse legumes dependson their nutrient composition, ruminal and post-ruminal digestibility, and on the presenceof secondary compounds that may interact with the rumen microbial population, therebylimiting nutrient utilisation (
Although the fermentation pattern of forages can be estimated in vitro by gas production
techniques (there are concerns aboutusing this technique to evaluate the CP quality of tropical forages. The widely used rumenin situ technique to measure CP degradation may overestimate its degradability in dried andground leaves because small particles pass through the bag pores without being degradedalternative methods, such as in vitro incubationwith rumen fluid or with exogenous proteases () were developed to estimate ruminal degradation. However,application of gas production techniques to estimate ruminal CP degradation has not received much research attention. Further, the intestinal digestion of CP hasalso been assessed by incubation of rumen undigested residue with exogenous enzymes
The fermentation pattern and CP degradation of four tropical browse legumes of potential
interest as ruminant feeds were evaluated in vitro by a gas production technique. Results werecompared with other estimations using rumen fluid and proteases, and intestinal digestionof the CP fraction was evaluated in vitro. 2. Materials and methods
The browse legumes, chosen by the Instituto de Ciencia Animal (ICA, La Habana,
Cuba), were Acacia cornigera (ACA), Albizia lebbekoides (ALB), Enterolobium cyclo-carpum (ENT) and Leucaena leucocephala (LEU). Representative samples of leaves andsmall stems were harvested in March 2002 from the ICA arboretum, dried at 60 ◦C for
M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
48 h and ground to pass a 1 mm screen before being sent to the University of Zaragoza(Spain).
Microbial fermentation patterns were studied in vitro with a gas production technique
(in two incubation experiments. In each, 601 ± 0.54 mg of thebrowses were weighed into bottles as the only substrate (n = 3), or with 150 mg (n = 2) or300 mg (n = 2) of soluble corn starch (Sigma (S-4126), St. Louis, MO, USA). In addition,triplicate bottles of incubation solution without substrate were included as blanks. Rumencontents were obtained from two adult ruminally cannulated sheep just before their morningfeeding (1 kg of a 1:1 alfalfa hay:barley straw mixture), pooled, and used as inoculum at afinal proportion of 0.10 of total incubation volume. The anaerobic incubation solution wasmade by combining the ingredients under a stream of CO2 with the help of a light source(Bottles were filled with 80 ml of incubation solution, sealed andincubated at 39 ◦C. Pressure of gas produced in each bottle was recorded with a HD8804manometer with a TP804 pressure gauge (DELTA OHM, Italy) after 3, 6, 9, 12 and 24 h ofincubation. Readings were converted to volume by using a pre-established linear regressionbetween pressure recorded in the same type of bottles and known inoculated air volumes(n = 64; r = 0.996). Gas volume at each incubation time was expressed per unit of incubatedorganic matter (OM).
Incubations were stopped at 24 h by dipping the bottles in cold water and immediately
filtering their contents through sintered glass crucibles with a 100 m maximum pore size. A 5 ml sample of liquid filtrate was collected over 5 ml of 0.2N HCl and stored frozenuntil ammonia analysis. Solid residue was washed twice with warm water, dried at 60 ◦Cfor 48 h and stored until N determination. The N degradability (NDg) of each browse wasestimated for each incubation experiment as proposed by based on thelinear relationship between volume of gas produced and net ammonia accumulation whenadditional amounts of fermentable carbohydrate were added to the medium. The linearregression of volume of gas produced (x, ml) and the amount of ammonia (y, mg) after24 h, was extrapolated to the x intercept (i.e., ammonia released when there was no energyavailable and therefore no bacterial synthesis), and this was related to total incubated Nafter subtracting the ammonia contribution of the blank (i.e., without substrate) at time = 0. 2.2. Other in vitro estimates of N degradability
The N degradability was also estimated as the proportion of N disappearing after 24 h
from substrate added to two bottles of each browse, as the only substrate, also included inthe previously cited incubation trials (NDd; Section Duplicate samples of the browseswere incubated with proteases type XIV from Streptomyces griseus (Sigma (P-5147), St. Louis, MO, USA) in borate-phosphate buffer (NDe) according to as modified by Incubations were in a shaking water bath for24 h on two consecutive days, and duplicated blank tubes (i.e., without plant samples) wereused to correct the enzymatic contribution to residual N. M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–3502.3. In vitro intestinal digestion of crude protein
The intestinal digestibility of CP was determined according to the three-step proce-
dure of Pooled (n = 3) dry residue of legumes incubated for24 h in vitro in each experiment (Section were weighed into duplicate tubes ensuring15 mg N/tube (370–640 mg DM). Briefly, pooled residues were first digested in vitro withHCl–pepsin at pH 1.9 for 1 h, followed by pancreatin at pH 7.8 for 24 h. The reaction wasstopped by adding trichloroacetic acid, and the centrifugation supernatant was analysedfor N. Before calculation, the N contribution of the pepsin–pancreatin solution blank wassubtracted from soluble N. Digestibility was calculated as the proportion of total N that wassolubilised.
Dried plant samples were analysed for DM by drying at 105 ◦C for 24 h and OM by
ashing at 550 ◦C for 6 h. Kjeldahl N was determined in a Kjeltec 2300 Analyzer Unit (FossTecator, Sweden) using Se as the catalyst and 1% boric acid in titration. Neutral detergentfibre (NDF), acid detergent fibre (ADF), acid detergent lignin (ADL) and acid detergentinsoluble N (ADIN) were determined by methods of NDF, ADFand ADL are expressed exclusive of residual ash. Condensed tannins (CT) were extractedand analysed according to s modified by purified quebracho tannins (Roy Wilson Dickson Ltd., UK) as the standard. Total CT were considered to be the sum of free-CT and protein-binding and fibre-bindingCT. Residues of incubation were also analysed for N by the Kjeldahl method. Ammoniaconcentration was determined colorimetrically according to
Results were analyzed by ANOVA using STATISTIX 8
production from the browses was compared by time considering the average value of thebottles from each experiment to be the experimental unit. CP degradation was examined ina split plot design, considering species to be the main plot and method as the subplot. Whendifferences (i.e., P < 0.05) occurred, treatment means were compared by least significantdifference (P < 0.05), and Pearson’s correlations were used among N degradability methods. 3. Results
The NDF and ADL proportions rank the forages into two types (being ALB
and LEU with the highest, and ENT and ACA with the lowest, levels. ALB and ACAhad higher proportions of both free and total CT versus LEU, whereas ENT had a min-imal level. The ADIN relative to N was between 0.23 and 0.53, ranking the species as:ENT > LEU> ALB > ACA.
There were differences among species in the gas volume produced from fermentation
throughout the incubation period (P < 0.001), with that from ACA the highest, and
M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
Table 1Proportions (g/kg dry matter) of organic matter (OM), total N, neutral detergent fibre (NDF), acid detergent fibre(ADF), acid detergent lignin (ADL) acid detergent insoluble N (ADIN) and free and total condensed tannins (CT)of browse legumes
that from ALB the lowest. From 3 to 9 h, gas produced from ENT was higher than fromLEU (P < 0.05), but there were no differences from 12 h.
The pattern of gas volume produced, and ammonia N released, due to added starch in
both experiments was linear (P < 0.01), allowing regression equations between gas volumeproduced and the ammonia N concentration at 24 h of incubation to be established for incu-bation study in order to estimate N degradability (NDg) of the browses (Differences(P < 0.001) among browses in y intercepts represent potential degradation of browse CP. All equations had a high coefficient of determination and, within browses, equations in bothexperiments had similar coefficients. Ammonia N in the blanks for experiments 1 and 2were 14.3 and 11.5 mg.
Estimations of N degradability based on the gas/ammonia relationship (NDg) from equa-
tions for each experiment were compared with the other in vitro procedures, being disap-pearance of N after incubation with rumen fluid (NDd) and after incubation with fungalenzymes (NDe), as shown in there were clear differences in N degradability
Fig. 1. In vitro gas production pattern of A. cornigera (᭹), Albizia lebbekoides ( ), E. cyclocarpum ( ) and L. leucocephala ( ) (each point is the average of two incubation runs with three bottles per incubation; upper barsshow standard errors of means). M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
Table 2Equations established between volume of gas produced (x, ml) and the ammonia N (y, mg) after 24 h of in vitroincubation of samples without or with 150 or 300 mg starch, considering the two incubation experiments separately(n = 7) or together (n = 14)
y = 19.995 − 0.0775(±0.00198)xy = 19.331 − 0.0859(±0.00258)xy = 19.579 − 0.0814(±0.00840)xy = 13.786 − 0.1749(±0.00654)xy = 14.998 − 0.1537(±0.00430)xy = 14.183 − 0.1205(±0.01208)xy = 25.446 − 0.1018(±0.00633)xy = 25.152 − 0.1291(±0.00945)xy = 24.879 − 0.1094(±0.01573)xy = 20.624 − 0.1022(±0.00239)xy = 15.970 − 0.0751(±0.00242)xy = 18.262 − 0.0881(±0.01226)x
S.D.: standard deviation. Ammonia N of blanks at t = 0 for experiments 1 and 2 was 14.3 and 11.5 mg.
among browses (P < 0.01), ranking ENT > ACA, and LEU > ALB, a trend to method dif-ferences (P = 0.06) suggests that NDd gave higher estimations than NDg, with NDe havingintermediate values. However, the interaction of species × method (P = 0.03) indicates thatdifferences among methods only occurred for ENT and LEU. In addition, NDe values werelowest for ENT, whereas NDe gave higher coefficients than NDg for LEU (P < 0.05). Pear-son’s correlations among the three methods (P < 0.05) show coefficients of 0.94 (betweenNDg and NDd), 0.83 (NDg and NDe) and 0.94 (NDd and NDe).
In vitro intestinal digestion, either as a fraction of residual N after 24 h of in vitro
incubation with rumen inoculum, or as a proportion of total N of the browses, is in For comparison, total feed N and estimated net available N at the intestines (i.e., total Nminus the proportion of ADIN and NDd) are also listed. Both ACA and LEU had higher(P = 0.04) in vitro digestion of undegraded N after incubation with rumen inoculum, but
Table 3In vitro degradability of N (g/kg) of the browse legumes according to the three evaluation methods (i.e., ammoniaand gas relationship, NDg; disappearance after incubation in rumen fluid, NDd; incubation with proteases, NDe)
Standard error of means for the interaction of species × method = 0.0447 (n = 2). Within species, and for overallspecies and methods, different letters show differences among means (P < 0.05). M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
Table 4In vitro intestinal digestion of N from the browse legumes and their undegradable fractions (g/g N)
Different letters show differences among means in rows (P < 0.05).
b Feed N − (NDd + ADIN) as a proportion of feed N.
there were no differences between LEU and ENT. This affected enzymatic digestion of totalN, which was also higher for ACA and LEU versus ALB and ENT (P = 0.005). 4. Discussion
The pattern of in vitro microbial fermentation to 24 h of incubation in this study agrees
with the high CT content of ACA, it had the higher volumeof gas production which is consistent with its chemical composition. In contrast, ALBhad very low fermentation, probably because of both high lignification of NDF and itshigh level of CT. As shown by gas produced from ENT increasedrapidly to 12 h, but fermentation was minimal thereafter. Considering the low CT and lignincontent of E. cyclocarpum, this could be due to negative effects of saponins on rumen protozoa (and cellulolytic bacteria (
The need for a rapid, simple, precise and inexpensive method to evaluate forage N sources
as potential ruminant feeds is clear. Estimation of available N by subtraction of ADIN is notaccurate, since a variable proportion of this fraction is degraded (Although the in situ technique has been widely used for tropical forage evaluation, and isoften considered to be the reference method, the likelihood that some particles wash outwithout being fermented by microbes (and low microbialactivity due to the presence of secondary compounds cannot beovercome. In an unpublished study with samples of the tested species incubated in Dacronbags (45 m pore size) for 24 h in the rumen of cows, N degradability coefficients were0.397, 0.689 and 0.763 for ALB, ENT and LEU. These values are far from what wasobserved here with other techniques for ALB and LEU; however the major concern was thehigh magnitude of the washing losses, accounting for 0.17, 0.38 and 0.53 of degraded N,respectively. These results show that the usefulness of this technique could be considerablyrestricted by the high proportion of particles below the pore size mainly in ground driedleaves of some species.
One of the major constraints to accuracy of method of incubation of feeds with rumen
fluid is underestimation of N degradation due to microbial synthesis that increases the pro-portion of N in the incubation residue (This problem can
M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
be overcome by utilisation of microbial inhibitors and short incubation times but it is not suitable for slowly degradable N sources. Incubation with fungal pro-teases tends to give higher values than rumen fluid, but the response is substrate dependant
There are few literature references of N degradability estimations using the in vitro
gas/ammonia relationship (estimates of N degradability of cereal straws by thegas/ammonia relationship with those of other in vitro methods, and reported a correlationbetween the gas method and pepsin solubility, but not with the fungal proteases or buffer Nsolubility, and higher values with fungal proteases versus the gas/ammonia relationship. Inour case, NDd had similar or higher values than NDg, suggesting that microbial contam-ination of residues of plants high in secondary factors after 24 h does not seem to impactaccuracy of the N degradation estimates. Statistical comparison among methods to measuredegradability showed that differences only occurred for ENT and LEU, and the lower valueof NDe for ENT might be due to reduction of N solubility by saponins (thus underestimating degradability, whereas this seems not to be the case when incubatingwith rumen fluid (i.e., NDg and NDd). No effect of saponins from E. cyclocarpum on CPdegradability by rumen fluid occurred in vitro (Regardless, the threemethods were highly correlated, showing that all can be used to determine N degradabilityof these tropical legumes.
Residual variability within the NDg method was much higher (i.e., cv = 0.26) than for
NDd (0.14) and NDe (0.05), and also compared to although it waslower than in Whereas the former used highly degradable CPsources, the latter examined forages with low degradability and low N, which would in-crease the magnitude of error. According to observed varia-tion may be related to differences in composition of rumen inoculum, although in our casethe statistical analysis of results from NDg alone showed no differences between experi-ments. In any case, a larger number of incubation trials would be desirable to reduce thisvariability.
Knowledge of N utilisation in the intestines is key to accurate estimation of the true value
of tropical forages as CP sources for ruminants, since some of their secondary compoundsmay impair intestinal digestion. The in vitro method used here has been shown to be wellcorrelated with in vivo results and is sensitive to the presence ofcompounds that reduce intestinal protein utilisation (edintestinal digestibility coefficients of CP reaching the duodenum were low, probably dueto the presence of tannins, and the high proportion of ADIN which is not easily digestedby intestinal enzymes. This would be the case for ALB, with low intestinal digestibility ofits undegraded CP fraction, and the low value observed with ENT might be also be due toformation of slowly digested saponin–protein complexes (In any case,values are similar to previously examined tropical legumes (e.g., coefficients of these browse species are within the range reported by for browses. Comparison of coefficients among browses was similar relative to intestinaldigestion of CP, and values for ACA and LEU showed that almost all available CP wasdigested, whereas for ALB a substantial proportion of it was not. The bias for ENT, amongboth estimations, is because a proportion of its ADIN was degraded. M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–3505. Conclusions
The relationship between gas production and ammonia concentration in 24 h in vitro
incubation is a useful and inexpensive procedure to estimate microbial N degradability oftropical browse legumes, and avoids the bias caused by high particle losses of ruminal insitu incubations of dried and ground leaves. In addition, this technique has been shown tobe as good as other methods, such as N degradation with rumen fluid or fungal enzymes(but being highly correlated with them), although there are issues about possible bias ofthe estimates when condensed tannins or saponin rich feedstuffs are incubated with fungalenzymes.
Among the four browses examined, A. cornigera had the highest gas production, as-
sociated with both a moderate N degradability and a considerable intestinal digestibility,indicating its nutritive potential as a ruminant feed in the tropics. In contrast, the nutritivevalue of A. lebbekoides, as a source of both CP and ruminally fermentable energy, was low. References
Analytical Software, 2003. STATISTIX 8 for Windows. Analytical Software, Tallahassee, FL, USA. Aufr`ere, J., Graviou, D., Demarquilly, C., V´erit´e, R., Michalet-Doreau, B., Chapoutot, P., 1991. Predicting in situ
degradability of feed proteins in the rumen by two laboratory methods (solubility and enzymatic degradation). Anim. Feed Sci. Technol. 33, 97–116.
Broderick, G.A., 1987. Determination of protein degradation rates using a rumen in vitro system containing
inhibitors of microbial nitrogen metabolism. Br. J. Nutr. 58, 463–475.
Broderick, G.A., Yang, H.J., Koegel, R.G., 1993. Effect of steam heating alfalfa hay on utilization by lactating
dairy cows. J. Dairy Sci. 76, 165–174.
Butter, N.L., Dawson, J.M., Buttery, P.J., 1999. Effects of dietary tannins on ruminants. In: Cargill, J.C., Mueller-
Harvey, I. (Eds.), Secondary Plant Products. Antinutritional and Beneficial Actions in Animal Feeding. Not-tingham University Press, Nottingham, UK, pp. 51–70.
Calsamiglia, S., Stern, M.D., 1995. A three-step in vitro procedure for estimating intestinal digestion of protein
in ruminants. J. Anim. Sci. 73, 1459–1465.
Chaney, A.L., Marbach, E.P., 1962. Modified reagents for determination of urea and ammonia. Clin. Chem. 8,
de Boer, G., Murphy, J.J., Kennelly, J.J., 1987. Mobile nylon bag for estimating intestinal availability of rumen
undegradable protein. J. Dairy Sci. 70, 977–982.
Fondevila, M., Nogueira, J.C.M., Barrios Urdaneta, A., 2002. In vitro microbial fermentation and protein utilisation
of tropical forage legumes grown during the dry season. Anim. Feed Sci. Technol. 95, 1–14.
Francis, G., Kerem, Z., Makkar, H.P.S., Becker, K., 2003. The biological action of saponins in animal systems: a
Fukushima, R.S., Weimer, P.J., Kunz, D.A., 2002. Photocatalytic interaction of resazurin N-oxide with cysteine
optimizes preparation of anaerobic culture media. Anaerobe 8, 29–34.
Getachew, G., Makkar, H.P.S., Becker, K., 1998. The in vitro gas coupled with ammonia measurements for
evaluation of nitrogen degradability in low quality roughages using incubation medium of different bufferingcapacity. J. Sci. Food Agric. 77, 87–95.
Hess, H.D., Kreuzer, M., D´ıaz, T.E., Lascano, C.E., Carulla, J.E., Soliva, C.R., Machm¨uller, A., 2003. Saponin
rich tropical fruits affect fermentation and methanogenesis in faunated and defaunated rumen fluid. Anim. Feed Sci. Technol. 109, 79–94.
Humphreys, L.R., 1995. Diversity and productivity of tropical legumes. In: D’Mello, J.P.F., Devendra, C. (Eds.),
Tropical Legumes in Animal Nutrition. CAB International, Wallingford, UK, pp. 1–21.
Hvelplund, T., Weisbjerg, M.R., 1998. In vitro techniques to replace in vivo methods for estimating amino acid
supply. In: Deaville, E.R., Owen, E., Adesogan, A.T., Rymer, C., Huntington, J.A., Lawrence, T.L.J. (Eds.),
M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
In Vitro Techniques for Measuring Nutrient Supply to Ruminants, BSAS Occasional Publication No. 22, pp. 131–144.
Kaitho, R.J., Umunna, N.N., Nsahlai, I.V., Tamminga, S., van Bruchem, J., 1998. Digestibility of rumen undegrad-
able protein from 40 browse species measured by mobile bag and in vitro techniques. In: Deaville, E.R., Owen,E., Adesogan, A.T., Rymer, C., Huntington, J.A., Lawrence, T.L.J. (Eds.), In Vitro Techniques for MeasuringNutrient Supply to Ruminants, BSAS Occasional Publication No. 22, pp. 123–126.
Krishnamoorthy, U., Sniffen, C.J., Stern, M.D., Van Soest, P.J., 1983. Evaluation of a mathematical model of
rumen digestion and an in vitro simulation of rumen proteolysis to estimate the rumen-undegraded nitrogencontent of feedstuffs. Br. J. Nutr. 50, 555–568.
Krishnamoorthy, U., Steingass, H., Menke, K.H., 1990. The contribution of ammonia, amino acids and short
peptides to estimates of protein degradability in vitro. J. Anim. Physiol. Anim. Nutr. 63, 135–141.
Krishnamoorthy, U., Soller, H., Steingass, H., Menke, K.H., 1995. Energy and protein evaluation of tropical
feedstuffs for whole tract and ruminal digestion by chemical analysis and rumen inoculum studies in vitro. Anim. Feed Sci. Technol. 52, 177–188.
Licitra, G., Van Soest, P.J., Schadt, I., Carpino, S., Sniffen, C.I., 1999. Influence of the concentration of the protease
from Streptomyces griseus relative to ruminal protein degradability. Anim. Feed Sci. Technol. 77, 99–113.
Makkar, H.P.S., Blummel, M., Becker, K., 1995. In vitro effects of and interaction between tannins and saponins
and fate of tannins in the rumen. J. Sci. Food Agric. 69, 481–493.
Menke, K.H., Steingass, H., 1988. Estimation of the energetic feed value obtained from chemical analysis and gas
production using rumen fluid. Anim. Res. Dev. 28, 7–55.
Newbold, C.J., El-Hassan, S.M., Wang, J., Ortega, M.E., Wallace, R.J., 1997. Influence of foliage from African
multipurpose trees on activity of rumen protozoa and bacteria. Br. J. Nutr. 78, 237–249.
P´erez Maldonado, R.A., Norton, B.W., 1996. The effect of condensed tannins from Desmodium intortum and
Calliandria calothyrsus on protein and carbohydrate digestion in sheep and goats. Br. J. Nutr. 76, 515–533.
Raab, L., Cafantaris, B., Jilg, T., Menke, K.H., 1983. Rumen protein degradation and biosynthesis 1. A new method
of protein degradation in rumen fluid in vitro. Br. J. Nutr. 50, 569–582.
Rodr´ıguez, R., Mota, M., Fondevila, M., de la Fuente, G., 2005. In vitro fermentation of four tropical browse
legumes: estimation of the effect of tannins by gas production. In: Sandoval-Castro, C.J., Hovell, F.D.DeB.,Torres-Acosta, J.F.J., Ayala-Burgos, A.J. (Eds.), Herbivores—The Assessment of Intake, Digestibility andthe Roles of Secondary Compounds, BSAS Occasional Publication No. 34, Nottingham University Press,Nottingham, UK.
Siaw, D.E.K.A., Osuji, P.O., Nsahlai, I.V., 1993. Evaluation of multipurpose tree germplasm: the use of gas
production and rumen degradation characteristics. J. Agric. Sci. Camb. 120, 319–330.
Solanas, E., Castrillo, C., Balcells, J., Fondevila, M., Guada, J.A., 2003. Estimaci´on de la digestibilidad intestinal
de la prote´ına no degradada en rumen de diferentes suplementos proteicos mediante m´etodos in situ e in vitro. ITEA 24 (II), 696–698.
Terrill, T.H., Rowan, A.M., Douglas, G.B., Barry, T.N., 1992. Determination of extractable and bound condensed
tannin concentration in forage plants, protein concentrate meals and cereal grains. J. Sci. Food Agric. 58,321–329.
Theodorou, K.M., Williams, B.A., Dhanoa, M.S., McAllan, A.B., France, J., 1994. A simple gas production
method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol. 48, 185–197.
Van Soest, P.J., Robertson, J.B., Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber and nonstarch
polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597.
Wang, Y., McAllister, T.A., Yanke, L.J., Xu, Z.J., Cheeke, P.R., Cheng, K.J., 2000. In vitro effects of steroidal
saponins from Yucca schidigera extract on rumen microbial protein synthesis and ruminal fermentation. J. Sci. Food Agric. 80, 2114–2122.
Nitro_Science_White_Paper-4.28.13-EN-ALL Kyäni Nitro FXTM and Kyäni Nitro XtremeTM: The Science Behind Kyäni’s Nitric Oxide Precursors The statements and products referenced in this document and have not been evaluated by the United States Food and Drug Administration (FDA). The Kyäni products are not intended to diagnose, treat, cure or prevent any disease or condition. The information
Patient information The information in this leaflet is to guide your use of nortriptyline this medicine safely. Further information is available inside the What is nortriptyline for? Notriptyline is a medicine which may help improve your pain control. It is different from other pain relief drugs. It can help nerve PHARMACY DIRECTORATE pain, for example shooting or burning


 Evaluation of four tropical browse legumes as
nitrogen sources: Comparison of in vitro gas
M. Mota , R. Rodr´ıguez , E. Solanas , M. Fondevila
a Departamento de Producci´on Animal y Ciencia de los Alimentos, Universidad de Zaragoza,
Miguel Servet 177, 50013 Zaragoza, Spain
b Instituto de Ciencia Animal, Ctra. Central km 47.5, San Jos´e de las Lajas,
Abstract
Evaluation of four tropical browse legumes as
nitrogen sources: Comparison of in vitro gas
M. Mota , R. Rodr´ıguez , E. Solanas , M. Fondevila
a Departamento de Producci´on Animal y Ciencia de los Alimentos, Universidad de Zaragoza,
Miguel Servet 177, 50013 Zaragoza, Spain
b Instituto de Ciencia Animal, Ctra. Central km 47.5, San Jos´e de las Lajas,
Abstract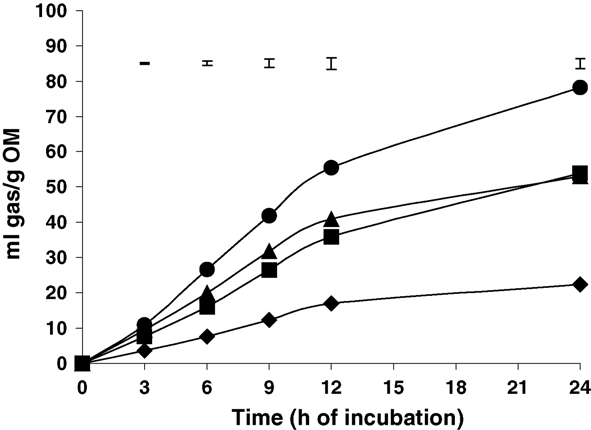 M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
Table 1Proportions (g/kg dry matter) of organic matter (OM), total N, neutral detergent fibre (NDF), acid detergent fibre(ADF), acid detergent lignin (ADL) acid detergent insoluble N (ADIN) and free and total condensed tannins (CT)of browse legumes
that from ALB the lowest. From 3 to 9 h, gas produced from ENT was higher than fromLEU (P < 0.05), but there were no differences from 12 h.
M. Mota et al. / Animal Feed Science and Technology 123–124 (2005) 341–350
Table 1Proportions (g/kg dry matter) of organic matter (OM), total N, neutral detergent fibre (NDF), acid detergent fibre(ADF), acid detergent lignin (ADL) acid detergent insoluble N (ADIN) and free and total condensed tannins (CT)of browse legumes
that from ALB the lowest. From 3 to 9 h, gas produced from ENT was higher than fromLEU (P < 0.05), but there were no differences from 12 h.