brief communications
rhesus or cynomolgus macaques successfullyyielded product from both batches. Sequenc-ing and phylogenetic analyses indicate that
those for CHAT 6039 were from cynomolgusmacaques (Fig. 1).
chimpanzee cellular components in two OPVCHAT stocks, together with the positive iden-
sequences, provides no support for thehypothesis that these materials were responsi-
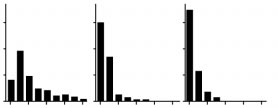
ble for the entry of HIV into humans and the source of AIDS. N. Berry*, C. Davis*, A. Jenkins*, D. Wood†, P. Minor†, G. Schild‡, M. Bottiger§, H. Holmes*, N. Almond* Divisions of *Retrovirology and †Virology, Figure 1 Maximum-likelihood phylogenies were estimated for three V3–V5 data sets: HIV-1 sequences from the Democratic Republic of Congo
‡National Institute for Biological Standards and
(423 base pairs), global isolates (426 base pairs), and Congo and global isolates combined (396 base pairs). Given a phylogeny with tips
Control, Blanche Lane, South Mimms, Potters Bar,
labelled according to subtype, the subtype diversity ratio (SDR) was calculated as the mean path length between tips of the same subtype divid-
ed by the mean path length between tips of different subtypes. For the phylogeny of the global isolates, 11 subtypes were allocated according
to standard HIV-1 nomenclature7. For the Congo phylogeny, 11 subtypes were allocated so as to minimize the SDR score, using a heuristic opti-
§Karolinska Institute, 17177 Stockholm, Sweden
mization algorithm. This assignment is that which gives the maximum possible subtype structure for the Congo phylogeny. The global phyloge-
1. Hooper, E. The River: A Journey to the Source of HIV and AIDS
ny gave an SDR of 0.33 and the Congo a value of 0.57. The analysis was repeated after removal of the Congo and global sequences previously
identified as intersubtype recombinants4,5. Our analysis will only be affected if recombination breakpoints fall within the V3–V5 region, so
2. Garrett, A. J., Dunham, A. & Wood, D. The Lancet 342, 932–933
excluding recombinants changes the SDR only marginally (0.35 for the global phylogeny; 0.58 for the Congo). SDR values were similar when
3. Dickson, D. Nature 407, 117 (2000).
Congo isolates were assigned to different numbers of subtypes (for example, 0.59 and 0.55 in the case of 8 and 14 subtypes, respectively). To
4. Hahn, B., Shaw, G., DeCock, K. & Sharp, P. Science 287, 607–614
assess the significance of the difference between the global and Congo SDRs, we obtained a null distribution by simulating phylogenies under
an exponential growth coalescent process inferred from env gene sequences of subtype A (ref. 6), which is common in Africa. The frequency
5. Berry, N. J. et al. J. Hum. Virol. 1, 457–468 (1998).
distribution of minimum SDR values for these simulated phylogenies is shown in blue. Inset: normalized frequency distributions of intrasubtype
6. WHO Network for HIV Isolation and Characterisation AIDS Res.Hum. Retrovir. 10, 1327–1343 (1994).
path lengths (above the line) and intersubtype path lengths (below the line), plotted on the same horizontal scale (0.0–0.8 substitutions per site),
7. Zhu, T. et al. Nature 391, 594–597 (1998).
for the global and Congo phylogenies. See supplementary information for details of trees and phylogenetic methods.
before the vaccination campaign2, supporting
mean between-subtype pairwise distance. Human immunodeficiency virus
a model of ‘natural transfer’ from chim-
Phylogeny and the
panzees to humans3. If this timescale is cor-
to subtypes by their phylogenetic relationship
rect, then the OPV theory remains a viable
to global strains, we used a heuristic algo-
origin of HIV-1
hypothesis of HIV-1 origins only if the sub-
rithm to assign subtypes such that the sub-
The origin of human immunodeficiency panzees before their transmission to humans. Congo and global phylogenies differ signifi-
virus type 1 (HIV-1) is controversial. We
It has been suggested that the distinctive
cantly in the SDR statistic, with the former
structure of the global group-M tree, which
the Democratic Republic of Congo in Africa
has been called a ‘starburst’ because of the
phylogenetic trees simulated under a model
have a quantitatively different phylogenetic
apparently simultaneous appearance of viral
of exponential population growth6 (Fig. 1; see
tree structure from those sampled in other
subtypes, is consistent with the transfer of
supplementary information). This result is
parts of the world. This indicates that the
multiple viral lineages from chimpanzees to
conservative because the minimum possible
structure of HIV-1 phylogenies is the result of
humans1. To test this, we analysed partial env
ratio value (representing maximum subtype
sequences (V3–V5) of 197 HIV-1 isolates
structure) was used in the Congo analysis.
human populations alone, and is not due to
sampled in 1997 from the Congo4, a likely
Furthermore, although subtypes can be clear-
multiple cross-species transmission initiated
ly identified in the distribution of pairwise
distances for the global sequences (Fig. 1,
inset), there is much less distinction between
plus 223 sequences representing the global
intra- and intersubtype comparisons for the
HIV-1 (the viruses responsible for the major-
diversity of HIV-1 (including all known sub-
ity of global AIDS cases) emerged as a result
types), reveals comparable genetic diversity in
domly chosen Congo sequences, it is difficult
of the vaccination of about one million peo-
ple, who were largely living in the Congo
strains, with many Congo lineages falling
belong to the same or to different subtypes.
from 1957–60, with an oral vaccine against
basal to the origin of each subtype as current-
polio virus that had allegedly been cultured in
ly defined by the phylogeny of global strains5.
global phylogenies probably result from dif-
chimpanzee kidneys1. This is claimed to have
We tested whether the structure of the Congo
ferent epidemiological histories. As many
phylogeny differed from that of the global
Congo strains appear to be basal, we propose
panzee simian immunodeficiency virus, the
that each global subtype is the result of the
diversity ratio (SDR) of the two phylogenies
chance exportation of some Congo strains to
Conversely, phylogenetic analysis of HIV-1
(Fig.1). This is defined as the ratio of the
other geographical regions, thus producing an
sequences indicates that group M originated
mean within-subtype pairwise distance to the
apparent starburst. Such founder effects have
NATURE | VOL 410 | 26 APRIL 2001 | www.nature.com
2001 Macmillan Magazines Ltd brief communications
been proposed to explain the phylogenetically
to return to nursing also revert to an arrhyth-
distinct subtypes B and E of HIV-1 group M
(ref. 2). The observation that many Congo
strains fall basal to the global subtypes also
each composed initially of 2,000–2,500 for-
suggests that previous phylogenetic analysis
agers, their queen and young (sib) brood.
has underestimated the number of lineages
that pre-date 1957–60, and hence underesti-
known to induce behavioural reversion8 , and
mated the minimum number of cross-species
indeed the division of labour was reorganized
transmissions necessary to reconcile the OPV
in these colonies: many bees continued to
forage, participating in little or no nursing
behaviour; some foragers reverted to nursing
the Congo are evidence that the claim of the
and stopped foraging completely, or almost
OPV theory1 that it is “probably the only
hypothesis of origin that can readily explain
As in typical colonies with young nurses6,
the starburst phenomenon” is incorrect. Our
brood care in our experimental colonies was
results give us no reason to doubt that the last
performed around the clock, with no diurnal
oscillations (Fig. 1b). The uninterrupted
nursing occurred because individual bees had
Andrew Rambaut*, David L. Robertson*,
reverted to arrhythmic activity: analysis of
Oliver G. Pybus*, Martine Peeters†, Edward
individually tagged reverted nurses (nǃ66)
C. Holmes*
revealed that brood care was performed by
*Department of Zoology, University of Oxford, South
arrhythmic bees nursing day and night, rather
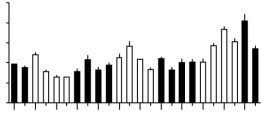
Figure 1 Reverted nurses care for brood with no diurnal rhythm.
than by rhythmic bees working in shifts (Fig. e-mail: [email protected]
Brood care was observed under dim red light (invisible to bees7)
1c). We found that reversion also affected the
†Laboratoire Retrovirus, IRD, BP 5045, 34032
every 3 h for three days. Observations of brood care6: six 10-min
activity–rest cycle: 21 reverted bees (31.8%)
visual scans of individually tagged bees in the vicinity of the brood.
cared for the brood in seven or more consecu-
1. Hooper, E. The River: A Journey to the Source of HIV and AIDS
Foraging observations were made as described6. a, Reorganization
tive observations for 21 hours or longer; for-
of division of labour in reversion colonies: frequency distributions of
agers, in contrast, rest daily for periods of
2. Korber, B. et al. Science 288, 1789–1796 (2000).
brood care differed significantly (chi-square test, P<0.05) for bees
3. Hahn, B., Shaw, G., de Cock, K. M. & Sharp, P. M. Science 287,
never observed foraging (0, left plot), observed foraging once (1,
4. Vidal, N. et al. J. Virol. 74, 10498–10507 (2000).
middle), or observed foraging more than once (>1, right). b, Colo-
basis of this striking natural behavioural plas-
5. Los Alamos National Laboratory HIV Sequence Database,
nial analysis. Mean (DŽs.e.) number of brood care events per obser-
ticity is unknown. There may be task-depen-
vation period during the day (white bars) and night (black bars) (nǃ6
dent changes in a central clock mechanism,
6. Pybus, O. G., Rambaut, A. & Harvey, P. H. Genetics 155,
scans per observation). To test comprehensively for diurnal rhythms,
7. Robertson, D. L. et al. Science 288, 55–57 (2000).
we pooled the data into two half-day categories and compared the
clock, or an effect resulting from nursing
Supplementary information is available on Nature’s website at
amount of brood-care activity between them; this analysis was
behaviour that overrides the clock output.
www.nature.com or as paper copy from the London editorial office of
repeated for eight different half-day combinations. No behavioural
Comparing these possibilities should help to
rhythms were detected (P>0.05, chi-square tests with Bonferonni
clarify the nature of the cellular and molecu-
correction). Results were similar for two other colonies (data not
lar4 bases of chronobiological plasticity.
shown). Foragers and reverted nurses did not differ in age
Chronobiology
(29.9DŽ0.2 days, nǃ26, and 29.8DŽ0.4, nǃ8, respectively;
brood to maturity in all three colonies. Reversal of honeybee
Pǃ0.76, unpaired t-test). c, Individual analyses. Number of scans
Although we did not test other possible con-
with brood care (days double-plotted). Bars at the bottom right show
behavioural rhythms
the light–dark regime outside: black, night; white, day). Sixty-six
have wider implications, given the conserva-
reverted nurses were analysed individually. Y44: example of a bee
tion of some molecular components of bio-
Adult honeybees have sleep-like states1,2 active around the clock and showing no diurnal rhythm in brood care logical clocks10and of sleep regulation11,12.
(P>0.05; statistical analyses as above); this behaviour was seen in
Guy Bloch*, Gene E. Robinson*†
80.3% of reverted nurses. Y4: example of a bee active around the
*Department of Entomology, †Neuroscience Program,
rhythms as they mature4,5. But whereas dis-
clock and with a weak diurnal rhythm in brood care (P>0.05); this
University of Illinois, 505 South Goodwin Avenue,
ruption of our sleep cycles and synchronized
behaviour was seen in 15.2% of reverted nurses. W53: one of only
internal rhythms may adversely affect our
three bees (4.5%) showing clear diurnal rhythms (P<0.05).
physiology and performance3, we show here
1. Kaiser, W. & Steiner-Kaiser, J. Nature 301, 707–709 (1983).
that honeybees can revert to certain arrhyth-
nal circadian clock for timing visits to flowers
2. Kaiser, W. J. Comp. Physiol. 163, 565–584 (1988).
3. Turek, F.W. & Zee, P. C. Regulation of Sleep and CircadianRhythms (Dekker, New York, 1999).
knowledge, this chronobiological plasticity is
4. Toma, D. P., Bloch, G., Moore, D. & Robinson, G.E. Proc. Natl
the first example in any animal of a socially
their behavioural development, with their
Acad. Sci. USA 97, 6914–6919 (2000).
mediated reversal in activity rhythms.
hive-to-field transition being accelerated,
5. Stussi, T. & Harmelin, M. L. C. R. Acad. Sci. Hebd. Seances. Acad.Sci. D 262, 2066–2069 (1966).
6. Moore, D., Angel, J. E., Cheeseman, I. M., Fahrbach, S. E. &
changing colony conditions8. We therefore
Robinson, G. E. Behav. Ecol. Sociobiol. 43, 147–160 (1998).
iour development process that underlies the
investigated whether this plasticity extends to
7. Von Firsch, K. The Dance Language and Orientation of Bees
colony’s division of labour. Larvae must be
the bees’ behavioural rhythms, focusing on
(Harvard University Press, Cambridge, 1967).
8. Huang, Z. Y. & Robinson, G. E. Behav. Ecol. Sociobiol. 39, 147–158
fed around the clock and are ‘nursed’ in the
the reversion from foraging to nursing as a
hive by young bees (5–15 days old) that work
9. Schulz, D. J. & Robinson, G. E. J. Comp. Physiol. A 184, 481–488
without any overt behavioural rhythms6. At
clock. This reversion occurs in response to a
severe shortage of nurse bees and is associated
10. Dunlap, J. C. Cell 96, 271–290 (1999). 11. Shaw, P. J., Cirelli, C., Greenspan, R. J. & Tononi, G. Science 287,
begins to forage outside the hive for pollen
with changes in exocrine, endocrine and neu-
and nectar, an activity that calls for an inter-
rochemical processes8,9. Do foragers induced
12. Hendricks, J. C. et al. Neuron 25, 129–138 (2000). 2001 Macmillan Magazines Ltd
NATURE | VOL 410 | 26 APRIL 2001 | www.nature.com
Name: ……………………… Group: ………. Date: …………… STEROIDS, PROSTAGLANDINS AND TERPENS Topics to prepare for the classes: 1. Structure, classification and biological functions of terpens and terpenoids. p. 1177- 2. Structure and biological functions of steroids. p. 1173-1176 3. Structure and biological functions of prostaglandins. p. 1176 - 1177 Problem 1
Arch. Environ. Contam. Toxicol. 48, 242–250 (2005)DOI: 10.1007/s00244-003-0262-7Comparative Sublethal Toxicity of Nine Pesticides on Olfactory LearningPerformances of the Honeybee Apis melliferaA. Decourtye,1 J. Devillers,2 E. Genecque,3 K. Le Menach,4 H. Budzinski,4 S. Cluzeau,1 M. H. Pham-Delgue31 Association de Coordination Technique Agricole, Maison des Agriculteurs, La Tour de Salvag
 brief communications
brief communications