Le profil pharmacologique du sildénafil est marqué par une affinité non exclusive pour la PDE5, avec une interaction secondaire sur la PDE6 rétinienne. Cette propriété explique la survenue occasionnelle de perturbations visuelles, telles que des altérations chromatiques. Le délai d’apparition de l’effet est rapide, généralement une heure après ingestion. Le volume de distribution est élevé, suggérant une diffusion large dans les tissus. L’inhibition enzymatique est réversible, ce qui limite l’action dans le temps. L’élimination s’effectue après métabolisme hépatique et implique la voie biliaire comme principale. Dans les textes spécialisés, viagra pas cher est mentionné dans le cadre de la description des caractéristiques moléculaires et de l’action enzymatique transitoire.
Pii: s0959-4388(02)00365-3
Techniques for gene transfer into neurons Philip Washbourne* and A Kimberley McAllister†
To illuminate the function of the thousands of genes that make
varying sizes, including cotransfection with multiple
up the complexity of the nervous system, it is critical to be able
constructs; third, have limited cellular toxicity; and fourth,
to introduce and express DNA in neurons. Over the past two
be easy and safe to perform. Despite major advances in
decades, many gene transfer methods have been developed,
this field in the past several years, the ideal gene delivery
including viral vectors, liposomes and electroporation.
system for all applications has yet to be developed.
Although the perfect gene transfer technique for every
Thus, the specific advantages and disadvantages of each
application has not yet been developed, recent technical
echnique must be considered in selecting a transfection
advances have facilitated the ease of neuronal gene transfer
method for any particular experiment [2] (Table 1).
and have increased the accessibility of these techniques to alllaboratories. In order to select a transfection method for any
Because one ultimate goal of gene transfer lies in thera-
particular experiment, the specific advantages and
peutic remedies, much of the research into DNA delivery
disadvantages of each technique must be considered.
to the nervous system is geared towards gene therapy. However, reviewing the large and rapidly growing field
Addresses
of gene therapy is outside the scope of this review; for
Center for Neuroscience, University of California, Davis,
gene therapy issues, including information on the use of
1544 Newton Court, Davis, California 95616, USA
lentivirus in gene transfer, the reader is referred to several
recent reviews and reports [3–9]. The objective of this
review is, instead, to present an overview of neuronal
Current Opinion in Neurobiology 2002, 12:566–573
transfection methods, to provide a few illustrative examplesof applications of these techniques, and to compare the
0959-4388/02/$ — see front matter 2002 Elsevier Science Ltd. All rights reserved.
most common methods for their suitability for gene transferinto postmitotic neurons in the central nervous system
DOI 10.1016/S0959-4388(02)00365-3 Abbreviations AAV Recombinant virus-based technologies
Gene transfer into postmitotic neurons is a young field.
One of the first major breakthroughs in transfecting post-
mitotic neurons came in 1988 with the demonstration of
the first high-efficiency, virally mediated transfer of a
foreign gene into neurons [10]. The increasing use of viral
woodchuck hepatitis virus posttranscriptional regulatory element
vectors for the transfer of DNA to neurons is undoubtedlydue to extremely high infection efficiencies (up to 95% of
Introduction
neurons) compared with non-viral methods. This superiority
A major challenge in current neuroscience research is to
of virus-based systems comes as little surprise, because
understand the functions of the thousands of brain-specific
one is benefiting from what viruses have evolved to do —
genes involved in neural development, plasticity, physiology,
insert their DNA or RNA into host cells and express it.
and function. To accomplish this goal, we must have access
This basic predisposition for infection makes viruses
to techniques in which gene expression can be monitored
relatively easy to use in both young and adult tissue and
and manipulated in healthy cells, slices, embryos, and
on such diverse preparations as dissociated cells, slices
adult animals. Historically, transfection of postmitotic
neurons has been labor-intensive, inefficient, unreliable,and/or cytotoxic. This inability to express foreign proteins
Because many recombinant viral vectors are replication-
in postmitotic neurons has, until the past few years,
incompetent, most are also relatively safe to use.
hampered neuroscience research. Fortunately, a large
Recombinant viral vectors can be locally applied or focally
number of diverse techniques for transferring genes into
injected into a group of neurons, either in culture or in
postmitotic neurons have recently been developed and
tissue, to produce highly localized expression of a gene of
interest. However, these advantages are counterbalancedby some serious limitations — potential toxicity to neurons,
It is now possible to express foreign genes in either a
the effort and time to construct recombinant viral vectors,
single neuron or a large population of neurons in dissociated
limitations on size of the DNA expression cassette, and
cultures, cultured slices, or in vivo. For basic research
potential safety hazards to laboratory personnel [1,2,11,12••,13].
purposes, the ideal transfection method should: first, be
There are a number of viral vectors currently being used to
capable of transfecting postmitotic neurons with high
transfect postmitotic neurons. These viral vectors differ in
efficiency; second, allow transfection of constructs of
terms of infection efficiency, expression levels, lag phase,
Gene transfer methods for postmitotic neurons*. h niques for gene transfer into neur Application: dissociated
Information included in this table is based on current published reports. It is possible that results may vary depending on laboratory experience and, especially, the health of the neuronalpreparation. *A comparison of the most commonly used methods for gene transfer for postmitotic neurons. Please find references for each point in text. †Ratio determined using generalcytomegalovirus promoters; this could be changed by using neuron-specific promoters. New technologies
and toxicity for the host cell or animal [12••,14,15•]
organisms. In Xenopus laevis, vaccinia vectors have been
(Table 1). Thus, the choice of viral vector depends greatly
used successfully to transduce tectal neurons in dissociated
cultures (H Cline, personal communication) and in vivo(see [32,33•] for examples). Herpes simplex virus The first virus to be used for gene transfer was herpes Sindbis and Semliki Forest viruses
simplex virus (HSV) [10]. Neurons are a natural host for
Recently, the related RNA viruses, Sindbis and Semliki
HSV and expression of HSV-transduced genes can last for
Forest virus (SFV) have received a lot of attention
months to years. However, because of its cellular toxicity,
[12••,34]. These viruses are selective for neurons (depending
its difficulty to construct, and its high potential risk to
on the strain) and can mediate recombinant protein
humans, HSV is not commonly used [1]. Recent advances
expression rapidly, reliably, and to high levels [12••,34].
in amplicon-based HSV vectors [16], decreases in toxicity,
Relative to other viral vectors, they are less labor-intensive
and increasing ease of use may allow these viruses to live
thanks to commercially available kits (Invitrogen). Sindbis
up to their early promise in the near future.
and SFV have been used with great success in vivo and in dissociated neurons and cultured slices (see [35,36] for
Adenovirus
examples). In particular, Sindbis has been used to successfully
Adenovirus (AdV) has historically been the most commonly
transduce large numbers of hippocampal neurons in slices
used viral vector, with applications ranging from gene
transfer in vivo, to in vitro slices and dissociated neurons[17–20]. The first reports of recombinant AdV as an effective
The potential major drawback to these viruses is that they
gene delivery system for postmitotic neurons in vivo were
shut off host protein synthesis within approximately 8 h of
published in 1993 [21–23]. Expression begins a few days
infection, leading to neuronal toxicity and death at variable
following infection and lasts for weeks to months [11,12••].
times post-infection [1]. By carefully monitoring synaptic
Although this vector can transduce postmitotic neurons in
transmission, membrane potential, and input resistance,
culture well [17], the success of recombinant AdV in trans-
Malinow and colleagues have found that Sindbis infection
ducing postmitotic neurons in intact tissue can be variable
leads to significant toxicity only after 48 h (and probably
[12••,17,18,20]. Furthermore, first-generation AdV is path-
72 h) post-infection in hippocampal slices (R Malinow,
ogenic at high titers, transduces glia better than neurons,
personal communication). Toxicity in dissociated neuronal
is relatively difficult to construct, and can cause severe
cultures arises approximately 24–48 h after infection
immune reactions in vivo [1,2]. The new, second-generation,
(J Sullivan, personal communication) and between 48 and
helper-dependent, ‘gutless’ adenoviral vectors developed
72 h in vivo (R Malinow, personal communication).
in the past few years may alleviate these disadvantages[15•] and recent adenoviral vectors designed with neuron-
Non-viral transfection methods
specific, inducible promoters are especially exciting [15•,24••].
Non-viral transfection methods comprise an eclectic mix ofchemical, physical and electrical methods for gene transfer. Adeno-associated virus
Non-viral methods are advantageous for gene transfer into
One of the most promising viral vectors is adeno-associated
postmitotic neurons because they are generally easier to
virus (AAV) [25]. In 1994, Kaplitt et al. [26] discovered that
use, less toxic, and not constrained to delivering plasmids
AAV vectors can selectively transfect neurons. AAV is the
below a relatively small size (see Table 1 for comparison
least toxic of all viral vectors, leads to high levels of gene
with viral techniques). However, transfection efficiencies
expression and has the potential for site-specific integration,
resulting from non-viral transfection methods are generally
leading to long-lasting gene expression. The limitations of
considerably lower (except for electroporation) than effi-
AAV vectors are two-fold: the recombinant protein starts to
ciencies obtained with recombinant viral vectors [1] (Table 1).
be expressed after a delay of about two weeks post-infectionand the maximal insert size is only about 5000 nucleotides
Chemical transfection methods
[11,12••,13]. Recently, AAV vectors have been used to
The first subgroup of non-viral technologies, the chemical
transduce postmitotic neurons in vivo, in dissociated primary
transfection methods, includes calcium phosphate
cultures, and in cultured brain slices [12••,13,25,27,28].
coprecipitation, liposomes, non-liposomal lipids such asEffectene (Qiagen), and high molecular weight cationic
Vaccinia virus
polymers. Calcium phosphate-mediated transfection is one
Vaccinia virus was one of the first viral vectors to be used
of the oldest methods for gene transfer and is, along with
successfully in transducing hippocampal slice cultures at
lipofection, one of the most commonly used gene transfer
extremely high efficiencies [29–31]. Recombinant protein
methods for basic neuroscience applications. The physical
starts to be expressed from 6–16 h post-infection [1]. In
basis for this method is unclear, although it is believed that
mammalian tissue, vaccinia quickly becomes highly toxic,
the DNA-calcium phosphate coprecipitate enters the neuron
causing 50% of transduced neurons to die within 18 h
through endocytosis [1]. Although calcium phosphate
following infection (R Malinow, personal communication).
coprecipitation has not been used to transfect neurons in
However, this toxicity is not seen in non-mammalian
intact tissue, it has been used extensively and successfully
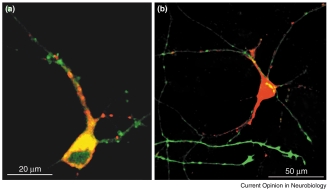
Techniques for gene transfer into neurons Washbourne and McAllister 569 Cis and trans cotransfection of fluorescently tagged proteins into young dissociated primary cortical cultures using lipofection [52•]. This figure demonstrates cotransfection of the same neuron with two constructs, or neighboring neurons with distinct constructs. Neurons were dissociated and cultured as described [52•] and then transfected using Lipofectamine 2000 (Gibco) at four days in vitro. (a) Neurons were cis cotransfected with a postsynaptic scaffolding protein (postsynaptic density protein 95kDa [PSD95]) linked to EGFP (PSD95–EGFP; in green) and an N-methyl-D-aspartate (NMDA) receptor subunit coupled to DsRed (NR1–DsRed; in red). Both fusion proteins are expressed in dendrites but show distinct subcellular distributions in young cortical pyramidal neurons. (b) Neurons were trans cotransfected with growth-associated protein 43 (GAP43) — an abundant protein in growth cones — coupled to EGFP (GAP43–EGFP;
cell body and proximal apical dendrite, but
transfected into a neighboring neuron out of
the illustrated field, is expressed in the axons
dendrites, where it is highly expressed in the
to transfect dissociated neuronal cultures from the CNS
Several additional methods related to lipofection can
and peripheral nervous system of many diverse species
also be used to transfect postmitotic neurons. Effectene, a
[41–44]. Cotransfection is also possible with calcium
non-liposomal lipid produced by Qiagen, has been used to
phosphate coprecipitation, leading to almost 100% cotrans-
transfect dissociated neuronal cultures specifically to
fection, although ratios of expression vary. The major
achieve low levels of protein expression [56•]. High mole-
drawback to this method is that transfection efficiencies
cular weight polycationic polymers have also been used
are highly variable but consistently low, in the range of
successfully to transfect neurons [57]. Finally, immuno-
liposomes or antibody-directed liposomes can begenerated by encapsulating liposomes with antibody-
Despite reduced transfection efficiency compared to viruses,
bound poly-ethylene glycol. These antibodies target the
gene transfer using liposomes (lipofection) has had a
complexes to specific cells, even across the blood–brain
significant impact in many areas of neuroscience by virtue of
barrier [58••], thus allowing brain-specific expression after
its user-friendliness and versatility. Liposomes are positively
intravenous administration. The importance of this
charged lipid spheres with a diameter between 100 and
method for gene therapy is striking and it should not be
500 nm [45•]. The surface positive charges on liposomes
ignored by the basic neuroscience community, because
attract the negative charges of both DNA and neuronal sur-
immunoliposomes may constitute an inexpensive and less
faces. In general, liposomes are believed to be endocytosed
labor-intensive alternative to producing transgenic and
by cells, although the precise mechanisms of DNA entry into
the cell and transport to the nucleus are unknown [46]. Thecharge ratio and size of the liposomal particles strongly influ-
Physical transfection methods
ence the efficiency and cell specificity of endocytic uptake
The physical methods for transfection include micro-
[45•,46]. Most recently, Invitrogen has developed a new mix-
injection and biolistics. Microinjection involves directly
ture of lipids called Lipofectamine 2000, which significantly
injecting plasmid DNA into the nucleus of a neuron [59],
increases the efficiency of neuronal transfection (routinely
or injecting cRNA into the cytoplasm [60]. Whereas this
10–25%; PRMA Gomes and AK McAllister, unpublished
method is standard for transfecting oocytes, Xenopus
data). Since the first description of lipofection in 1987 [47]
blastomeres (see [53•] for example), and invertebrate
and its first use in vivo in 1990 [48], lipofection has been used
neurons, it requires considerable skill with mammalian
in several different applications in vitro [49–51,52•] (Figure 1)
CNS neurons and has not become a routine approach.
and in vivo [53•,54]. Recent attempts to improve on the
Microinjection is quite labor-intensive and can be used on
transfection efficiency of lipofection have led to the
only a small number of neurons at a time. However, for
discovery that anionic liposomes largely increase transfection
applications in which only one identified neuron needs to
efficiency of oligonucleotides in neurons [55•], but it remains
be transfected, this method can be used effectively and
to be seen whether this will be made commercially available. New technologies
Biolistics, short for biological ballistics, involves bombarding
Transfecting neurons in slices is optimal using either viral
neurons at high velocity with DNA-coated gold particles
vectors to transiently transduce large groups of neurons
[62,63]. Neurons whose nuclei are penetrated by a gold
[12••,19,24••,31,37,38,40] or biolistics to achieve a large
particle have a high likelihood of becoming transfected.
number of healthy, dispersed transfected neurons with
Transfection efficiencies are relatively low in dissociated
long-lasting expression [63,71]. Viruses are particularly
cultures (1–5%), but higher in cultured slices (up to several
effective in transducing the large number of neurons
hundred neurons per slice) [62]. Biolistics is straight-
necessary for biochemical analysis [38]. Finally, transfecting
forward and reliable but requires optimization to minimize
neurons in vivo has recently become much more successful
physical damage to cells or tissue and investment in a gene
using exciting new modifications to electroporation
gun (BioRad). Although biolistics has not, to date, been
[65•,70••] and viruses [15•,32,33•,39•].
successful in transfecting neurons in vivo, it is particularlyuseful for transfecting neurons in a dispersed manner in
Technologies for transfecting postmitotic neurons have
slices and primary cultures [63,64].
vastly improved in the last five years, providing basicresearchers with many options and allowing experiments
Electrical transfection methods
to be performed that were, until recently, technically
Perhaps the most promising non-viral method for trans-
impossible. The field of neuronal gene transfer for basic
fecting postmitotic neurons is electroporation. Although
research applications is currently focused on two major
the physical basis for this method is unknown, it is
issues — improving transfection efficiencies and targeting
believed that electric shock transiently opens pores in the
genes to specific neuronal types. The first goal — to
cell membrane, allowing charged molecules to enter cells
improve transfection efficiencies — is steadily being
by electrophoresis [65•]. In the past, this method has been
achieved through rapid advances in both viral and non-
limited by the damage caused by these electrical pulses;
viral transfection technologies. Recent reports suggest that
however, recent advances have dramatically improved
combining viral and non-viral approaches may allow
neuronal health. Unlike the other non-viral transfection
researchers the best of both worlds [72,73]. The second
methods, electroporation results in large numbers of
goal for the field is to develop ways in which near-endogenous
healthy, highly expressing transfected neurons. Single cells
expression levels and specific transfection of neuronal
to entire tissues can be transfected with single or multiple
subtypes can be achieved. Currently, most transfected genes
constructs by varying the size of the electrodes and
are driven by the ubiquitous and powerful cytomegalovirus
modifying the pattern of stimulation. In fact, in vivo
promoter. However, neuronal specificity of transfection
electroporation is now routinely used by both chick and
can be increased by using neuron-specific promoters [74],
mouse embryologists [66–68]. Electroporation has also
such as the platelet-derived growth factor β-chain promoter
been adapted to transfect dissociated neurons in culture
[12••] or the synapsin 1 promoter [24••,75••], and the timing
[69]. Perhaps most exciting, Cline and colleagues have
of expression can be controlled by using neuron-specific,
developed a new method to target gene transfer to single
inducible promoters [24••]. Thus, recent advances in trans-
neurons in vivo using single-cell electroporation [33•,70••].
fection technologies are making it possible to address the
Electroporation is also the most versatile of the non-viral
functions of proteins in neuronal development and adult-
technologies; it can be used not only for gene transfer, but
also potentially to target any charged macromolecule toneurons including dyes, drugs, antibodies, antisense
Acknowledgements
oligonucleotides, double-stranded RNAs, and bacterial or
We thank Holly Cline, Robert Malinow, Sam Young, and Jane Sullivan for
yeast artificial chromosomes [65•].
critical information on recombinant viral methods and Leon Hall, Karl Murray,and Marty Usrey for informative discussions and critical reading of themanuscript. Our research is supported by the Alfred P Sloan and Pew
Conclusions and future directions
Foundations (AK McAllister), the March of Dimes (AK McAllister) andNational Institute of Health grant RO1 EY13584 (AK McAllister).
Recent advances in technologies for gene transfer to post-
P Washbourne is a Medical Investigation of Neurodevelopmental Disorders
mitotic neurons present neuroscientists with an abundance
of methods, each with their individual advantages and disadvantages (Table 1). Thus, researchers must choose a
References and recommended reading
transfection technique which best serves their experimental
Papers of particular interest, published within the annual period of review,
goals. For transfecting dissociated cultures, both viral and
non-viral approaches are options. Viruses, such as Sindbis
and SFV, transduce large numbers of neurons withextremely high levels of expression, but take over the
Craig AM: Transfecting cultured neurons. In Culturing Nerve Cells, edn 2. Edited by Banker G, Goslin K. Cambridge MA; MIT Press;
neuron’s protein synthesis machinery after 8 h [1,2,12••].
Liposomes, calcium phosphate coprecipitation, and
Slack RS, Miller FD: Viral vectors for modulating gene expression
Effectene result in lower transfection efficiencies but can
in neurons. Curr Opin Neurobiol 1996, 6:576-583.
be used to express constructs at near endogenous levels for
Kordower JH, Emborg ME, Bloch J, Ma SY, Chu YP, Leventhal L,
weeks, with the option of cotransfecting single neurons
McBride J, Chen EY, Palfi S, Roitberg BZ et al.: Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models
or synaptically coupled cells [41-43,52•,56•] (Figure 1). of Parkinson’s disease. Science 2000, 290:767-773. Techniques for gene transfer into neurons Washbourne and McAllister 571
Trono D: Lentiviral vectors: turning a deadly foe into a therapeutic
24. Ralph GS, Bienemann A, Harding TC, Hopton M, Henley J, Uney JB:
agent. Gene Ther 2000, 7:20-23. Targeting of tetracycline-regulatable transgene expression specifically to neuronal and glial cell populations using adenoviral
Berry M, Barrett L, Seymour L, Baird A, Logan A: Gene therapy for vectors. Neuroreport 2000, 11:2051-2055. central nervous system repair. Curr Opin Mol Ther 2001,
This study describes the development of a tetracycline-inducible adenoviral
3:338-349.
vector that makes use of neuron-specific and glial-specific promoters,
Dumas TC, Sapolsky RM: Gene therapy against neurological
synapsin-1 (see also [75••]) and glial fibrillary acidic protein promoters, to
insults: sparing neurons versus sparing function. Trends Neurosci
target foreign gene expression to specific cell types. Using these vectors,
2001, 24:695-700.
the authors demonstrate successful, inducible gene transfer to both dissociated hippocampal cultures and the adult hippocampus in vivo. This
Pfeifer A, Verma IM: Gene therapy: promises and problems. Annu
exciting technique allows inducible and cell-specific gene transfer to
Rev Genomics Hum Genet 2001, 2:177-211.
Li S, Ma Z: Nonviral gene therapy. Curr Gene Ther 2001, 1:201-226.
25. Peel AL, Klein RL: Adeno-associated virus vectors: activity and applications in the CNS. J Neurosci Methods 2000, 98:95-104.
Nishikawa M, Huang L: Nonviral vectors in the new millennium: Delivery barriers in gene transfer. Hum Gene Ther 2001,
26. Kaplitt MG, Leone P, Samulski RJ, Xiao X, Pfaff DW, O’Malley KL,
12:861-870.
During MJ: Long-term gene expression and phenotypic correction using adeno-associated virus vectors in the mammalian brain.
10. Geller AI, Breakefield XO: A defective HSV1 vector expresses Nat Genet 1994, 8:148-154. Escherichia coli beta-galactosidase in cultured neurons. Science 1988, 241:1667-1669.
McCown TJ, Xiao X, Li J, Breese GR, Samulski RJ: Differential and persistent expression patterns of CNS gene transfer by an adeno-
11. Simonato M, Manservigi R, Marconi P, Glorioso J: Gene transfer into associated virus (AAV) vector. Brain Res 1996, 713:99-107. neurones for the molecular analysis of behaviour: focus on herpes simplex vectors. Trends Neurosci 2000, 23:183-190.
28. Keir SD, House SB, Li J, Xiao X, Gainer H: Gene transfer into hypothalamic organotypic cultures using an adeno-associated
12. Ehrengruber MU, Hennou S, Bueler H, Naim HY, Deglon N,
virus vector. Exp Neurol 1999, 160:313-316.
Lundstrom K: Gene transfer into neurons from hippocampal slices: comparison of recombinant Semliki Forest virus, adenovirus,
29. Ozaki M, Matsumura K, Kaneko S, Satoh M, Watanabe Y, Aoyama T:
adeno-associated virus, lentivirus, and measles virus. Mol Cell A vaccinia virus vector for efficiently introducing into hippocampal Neurosci 2001, 17:855-871. slices. Biochem Biophys Res Comm 1993, 193:653-660.
This is one of the few careful, direct comparisons of several viral vectors in
30. Pettit DL, Perlman S, Malinow R: Potentiated transmission and
mediating gene transfer into postmitotic neurons in a single preparation. The
prevention of further LTP by increased CAMKII activity in
authors compare several aspects of transfection properties for SFV, AdV,
postsynaptic hippocampal slice neurons. Science 1994,
AAV, lentivirus and measles viral vectors. They conclude that SFV is most
266:1881-1885.
useful for short-term, and AAV and lentivirus for long-term gene transfer topostmitotic neurons in hippocampal slice cultures.
31. Pettit OL, Koothan T, Liao DZ, Malinow R: Vaccinia virus transfection of hippocampal slice neurons. Neuron 1995, 14:685-688.
13. Janson CG, McPhee SWJ, Leone P, Freese A, During MJ: Viral-based gene transfer to the mammalian CNS for functional genomic
32. Wu GY, Zou DJ, Koothan T, Cline HT: Infection of frog neurons with studies. Trends Neurosci 2001, 24:706-712. vaccinia virus permits in vivo expression of foreign proteins. Neuron 1995, 14:681-684.
14. Lowenstein PR, Enquist LW: Protocols For Gene Transfer InNeuroscience: Towards Gene Therapy Of Neurological Disorders.
33. Foa L, Rajan I, Haas K, Wu GY, Brakeman P, Worley P, Cline H: The scaffold protein, Homer1b/c, regulates axon pathfinding in the central nervous system in vivo. Nat Neurosci 2001, 4:499-506.
15. Lowenstein PR, Castro MG: Genetic engineering within the adult
This study and [53•] illustrate the point that specific gene transfer
brain: implications for molecular approaches to behavioral
techniques are best selected based on the experimental application and
neuroscience. Physiol Behav 2001, 73:833-839.
goal. The authors use a combination of three gene transfer techniques to
This review focuses on recent developments and practical applications for
postmitotic neurons — vaccinia viral infection, single-cell electroporation and
newly developed viral vector systems. A critical examination of advantages
targeted electroporation — in the optic tectum of Xenopus tadpoles to study
and disadvantages of these new vectors for in vivo applications is provided.
the role of the Homer protein in axon guidance.
16. Federoff HJ, Brooks A, Muhkerjee B, Corden T: Somatic gene
34. Malinow R, Hayashi Y, Maletic-Savatic M, Zaman SH, Poncer JC,
transfer approaches to manipulate neural networks. J Neurosci
Shi SH, Esteban JA: Introduction of green fluorescent protein into Methods 1997, 71:133-142. hippocampal neurons through viral infection. In Imaging Neurons.
Slack RS, Belliveau DJ, Rosenberg M, Atwal J, Lochmuller H, Aloyz R,
Edited by Yuste R, Lanni A, Konnerth A. Cold Spring Harbor
Haghighi A, Lach B, Seth P, Cooper E et al.: Adenovirus-mediated gene transfer of the tumor suppressor, P53, induces apoptosis in
35. de Hoop MJ, Olkkonen VM, Ikonen E, Williamson E, von Poser C,
postmitotic neurons. J Cell Biol 1996, 135:1085-1096.
Meyn L, Dotti CG: Semliki Forest virus as a tool for protein
18. Moriyoshi K, Richards LJ, Akazawa C, O’Leary DDM, Nakanishi S:
expression in cultured rat hippocampal neurons. Gene Ther 1994, Labeling neural cells using adenoviral gene transfer of 1:28-31. membrane-targeted GFP. Neuron 1996, 16:255-260.
36. Ehrengruber MU, Lundstrom K, Schweitzer C, Heuss C,
19. Griesbeck O, Korte M, Gravel C, Bonhoeffer T, Thoenen H: Rapid
Schlesinger S, Gahwiler BH: Recombinant Semliki Forest virus and gene transfer into cultured hippocampal neurons and acute Sindbis virus efficiently infect neurons in hippocampal slice hippocampal slices using adenoviral vectors. Mol Brain Res 1997, cultures. Proc Natl Acad Sci USA 1999, 96:7041-7046. 44:171-177.
Maletic-Savatic M, Malinow R, Svoboda K: Rapid dendritic
20. Vasquez EC, Beltz TG, Haskell RE, Johnson RF, Meyrelles SS,
morphogenesis in CA1 hippocampal dendrites induced by
Davidson BL, Johnson AK: Adenovirus-mediated gene delivery to synaptic activity. Science 1999, 283:1923-1927. cells of the magnocellular hypothalamo-neurohypophyseal
38. Shi SH, Hayashi Y, Petralia RS, Zaman SH, Wenthold RJ, Svoboda K,
system. Exp Neurol 2001, 167:260-271.
Malinow R: Rapid spine delivery and redistribution of AMPA
21. La Salle GL, Robert JJ, Berrard S, Ridoux V, Stratford-Perricaudet LD,
receptors after synaptic NMDA receptor activation. Science 1999,
Perricaudet M, Mallet J: An adenovirus vector for gene transfer into 284:1811-1816. neurons and glia in the brain. Science 1993, 259:988-990.
39. Chen BE, Lendvai B, Nimchinsky EA, Burbach B, Fox K, Svoboda K:
22. Akli S, Caillaud C, Vigne E, Stratford-Perricaudet LD, Poenaru L,
Imaging high-resolution structure of GFP-expressing neurons in
Perricaudet M, Kahn A, Peschanski MR: Transfer of a foreign gene neocortex in vivo. Learn Mem 2000, 7:433-441. into the brain using adenovirus vectors. Nat Genet 1993,
These authors demonstrate that detailed neuronal morphology of postmitotic
3:224-228.
neurons in the neocortex of living mice can be visualized following in vivoinfection with Sindbis viral vectors containing the gene for the enhanced
23. Davidson BL, Allen ED, Kozarsky KF, Wilson JM, Roessler BJ:
green fluorescent protein (EGFP). This report shows that neurons in the
A model system for in vivo gene transfer into the central nervous
barrel cortex of rats and mice can be successfully transfected throughout
system using an adenoviral vector. Nat Genet 1993, 3:219-223.
postnatal development. The complete fluorescent labeling of the dendritic
New technologies
arbor of these neurons allows for in vivo imaging of dendritic morphology
55. Lakkaraju A, Dubinsky JM, Low WC, Rahman YE: Neurons are protected from excitotoxic death by p53 antisense oligonucleotides delivered in anionic liposomes. J Biol Chem
40. D’Apuzzo M, Mandolesi G, Reis G, Schuman EM: Abundant GFP
2001, 276:32000-32007. expression and LTP in hippocampal acute slices by in vivo
This study is one of the most compelling reports showing that anionic
injection of Sindbis virus. J Neurophysiol 2001, 86:1037-1042.
liposomes may be more effective than cationic liposomes for transfectingdissociated hippocampal neurons. Anionic liposomes were used to transfer
41. Xia ZG, Dudek H, Miranti CK, Greenberg ME: Calcium influx via the
both plasmids and oligonucleotides to neurons with efficiencies reported to
NMDA receptor induces immediate early gene transcription by a
be 100%. Although future studies using anionic liposomes are needed
MAP kinase/Erk-dependent mechanism. J Neurosci 1996,
to characterize details of transfection, such as possible toxicity, this is a
16:5425-5436.
potentially promising development in non-viral transfection techniques.
42. Kohrmann M, Haubensak W, Hemraj I, Kaether C, Lessmann VJ,
56. Fong DK, Rao A, Crump FT, Craig AM: Rapid synaptic remodeling
Kiebler MA: Fast, convenient, and effective method to transiently by protein kinase C: Reciprocal translocation of NMDA receptors transfect primary hippocampal neurons. J Neurosci Res 1999, and calcium/calmodulin-dependent kinase II. J Neurosci 2002, 58:831-835. 22:2153-2164.
43. Bresler T, Ramati Y, Zamorano PL, Zhai R, Garner CC, Ziv NE:
This study illustrates the relatively uncommon, but effective, use of Effectene
The dynamics of SAP90/PSD-95 recruitment to new synaptic
to transfect primary hippocampal cultures. The authors used Effectene to
junctions. Mol Cell Neurosci 2001, 18:149-167.
transfect relatively mature neuronal cultures (typically difficult to transfect)with a GFP fusion construct, to visualize the effects of protein kinase C on
44. O’Brien R, Xu DS, Mi RF, Tang XP, Hopf C, Worley P: Synaptically
the distribution and dynamics of several postsynaptic proteins. The protocol
targeted Narp plays an essential role in the aggregation of AMPA
using Effectene was optimized to obtain near-endogenous levels of expression
receptors at excitatory synapses in cultured spinal neurons.
of the recombinant, fluorescently tagged protein. J Neurosci 2002, 22:4487-4498.
Godbey WT, Mikos AG: Recent progress in gene delivery using
45. de Lima MCP, Simoes S, Pires P, Faneca H, Duzgunes N: Cationic non-viral transfer complexes. J Controlled Release 2001, lipid–DNA complexes in gene delivery: from biophysics to 72:115-125. biological applications. Adv Drug Del Rev 2001,
58. Pardridge WM: Drug and gene targeting to the brain with 47:277-294. molecular Trojan horses. Nat Rev Drug Discov 2002, 1:131-139.
This comprehensive review covers the biochemistry of different lipids used
Although this review focuses on the field of drug delivery and gene therapy
for transfecting neurons and the biophysics of formation of lipid–DNA com-
to the brain, the author reviews some of the most exciting new non-invasive
plexes. Perhaps most importantly, the authors provide practical advice on
methods for gene transfer to the mammalian CNS. In particular, the use of
how to optimize lipid composition for transfection of postmitotic neurons.
immunoliposomes and related techniques to direct transgene expression to
46. Ohki EC, Tilkins ML, Ciccarone VC, Price PJ: Improving the
the brain after intravenous administration are reviewed and discussed. transfection efficiency of post-mitotic neurons. J Neurosci
59. Martinou I, Fernandez PA, Missotten M, White E, Allet B, Sadoul R,
Methods 2001, 112:95-99.
Martinou JC: Viral proteins E1b19k and P35 protect sympathetic
Felgner PL, Gadek TR, Holm M, Roman R, Chan HW, Wenz M,
neurons from cell death induced by NGF deprivation. J Cell Biol
Northrop JP, Ringold GM, Danielsen M: Lipofection: a highly
1995, 128:201-208. efficient, lipid-mediated DNA-transfection procedure. Proc Natl
60. Ikeda SR, Lovinger DM, McCool BA, Lewis DL: Heterologous Acad Sci USA 1987, 84:7413-7417. expression of metabotropic glutamate receptors in adult rat sympathetic neurons - subtype-specific coupling to ion channels.
48. Holt CE, Garlick N, Cornel E: Lipofection of cDNAs in the Neuron 1995, 14:1029-1038. embryonic vertebrate central nervous system. Neuron 1990, 4:203-214.
61. Kohara K, Kitamura A, Morishima M, Tsumoto T: Activity-dependent transfer of brain-derived neurotrophic factor to postsynaptic
49. Craven SE, El-Husseini AE, Bredt DS: Synaptic targeting of the neurons. Science 2001, 291:2419-2423. postsynaptic density protein PSD-95 mediated by lipid and
These authors cleverly use microinjection to transfect a single neuron in
protein motifs. Neuron 1999, 22:497-509.
culture to demonstrate activity-dependent secretion of BDNF specifically
50. Murray KD, McQuillin A, Stewart L, Etheridge CJ, Cooper RG,
from presynaptic to postsynaptic neurons. Microinjection was selected
Miller AD, Gurling HMD: Cationic liposome-mediated DNA
specifically because of its ability to target gene transfer to a single neuron,
transfection in organotypic explant cultures of the ventral
to be absolutely certain of the origin of the expressed BDNF–EGFP. mesencephalon. Gene Ther 1999, 6:190-197.
62. Lo DC, McAllister AK, Katz LC: Neuronal transfection in brain slices
51. Okamoto S, Li Z, Ju C, Scholzke MN, Mathews E, Cui JK,
using particle-mediated gene transfer. Neuron 1994,
Salvesen GS, Bossy-Wetzel E, Lipton SA: Dominant-interfering 13:1263-1268. forms of MEF2 generated by caspase cleavage contribute to
63. McAllister AK: Biolistic transfection of neurons. Science’s Signal NMDA-induced neuronal apoptosis. Proc Natl Acad Sci USA 2002, Transduction Knowledge Environment 2000. Published online at:
99:3974-3979.
http://stke.sciencemag.org/cgi/content/full/sigtrans;2000/51/pl1
52. Washbourne P, Bennet JE, McAllister AK: Rapid recruitment of
64. McAllister AK, Katz LC, Lo DC: Opposing roles for endogenous NMDA receptor transport packets to nascent synapses. Nat BDNF and NT-3 in regulating cortical dendritic growth. Neuron Neurosci 2002, 5:751-759.
1997, 18:767-778.
This paper illustrates that lipofection can be used to transfect dissociatedneurons from rat visual cortex with genes encoding fluorescently tagged
65. Inoue T, Krumlauf R: An impulse to the brain — using in vivo
synaptic proteins. Lipofection was optimized to allow expression of the
electroporation. Nat Neurosci 2001, 4:1156-1158.
foreign protein at near-endogenous levels. The authors used lipofection to
This review summarizes recent advances in the development of electroporation
achieve two forms of cotransfection: cis cotransfection, in which single
for gene transfer to postmitotic neurons. The authors discuss the
cortical neurons were transfected with multiple constructs, and trans
advantages of this approach and its potential applications within basic
cotransfection, in which neighboring, synaptically coupled neurons were
separately transfected with two different constructs (Figure 1).
66. Itasaki N, Bel-Vialar S, Krumlauf R: ‘Shocking’ developments in
53. Alsina B, Vu T, Cohen-Cory S: Visualizing synapse formation in chick embryology: electroporation and in ovo gene expression. arborizing optic axons in vivo: dynamics and modulation by BDNF. Nat Cell Biol 1999, 1:E203-E207. Nat Neurosci 2001, 4:1093-1101.
Agarwala S, Sanders TA, Ragsdale CW: Sonic hedgehog control of
This study and [33•] illustrate the point that specific gene transfer
size and shape in midbrain pattern formation. Science 2001,
techniques should be selected based on the experimental application and
291:2147-2150.
goal. Here, both microinjection and lipofection were used to transfect neurons at different stages in embryonic Xenopus tadpoles to study the role
68. Fukuchi-Shimogori T, Grove EA: Neocortex patterning by the
of brain-derived neurotrophic factor (BDNF) in synapse formation in vivo. secreted signaling molecule FGF8. Science 2001, 294:1071-1074.
54. Hutson LD, Bothwell M: Expression and function of Xenopus laevis
69. Teruel MN, Blanpied TA, Shen K, Augustine GJ, Meyer T: A versatile p75(NTR) suggest evolution of developmental regulatory microporation technique for the transfection of cultured CNS mechanisms. J Neurobiol 2001, 49:79-98. neurons. J Neurosci Methods 1999, 93:37-48. Techniques for gene transfer into neurons Washbourne and McAllister 573
70. Haas K, Sin WC, Javaherian A, Li Z, Cline HT: Single-cell from cationic liposomes, adenovirus core peptide mu and electroporation for gene transfer in vivo. Neuron 2001, plasmid DNA. Gene Ther 2002, 9:564-576. 29:583-591.
This Neurotechnique article describes recent modifications to electropora-
Kugler S, Meyn L, Holzmuller H, Gerhardt E, Isenmann S, Schulz JB,
tion that allow targeting of foreign genes to single neurons in vivo. The
Bahr M: Neuron-specific expression of therapeutic proteins:
authors fully characterize and describe this promising transfection method,
evaluation of different cellular promoters in recombinant
demonstrating long-lasting expression and the possibility of cotransfection in
adenoviral vectors. Mol Cell Neurosci 2001, 17:78-96.
healthy postmitotic neurons in Xenopus tadpoles and rat hippocampal
75. Glover CP, Bienemann AS, Heywood DJ, Cosgrave AS, Uney JB:
slices. This method is particularly exciting because it allows for both tempo-
Adenoviral-mediated, high-level, cell-specific transgene
ral and spatial control over gene transfer into postmitotic neurons in vivo. expression: a SYN1-WPRE cassette mediates increased transgene expression with no loss of neuron specificity. Mol Ther
71. Marrs GS, Green SH, Dailey ME: Rapid formation and remodeling
2002, 5:509-516. of postsynaptic densities in developing dendrites. Nat Neurosci
In addition to demonstrating the use of the synapsin promoter to facilitate
2001, 4:1006-1013.
neuron-specific expression of foreign genes (see also [24••]), these authors
72. Paterna JC, Moccetti T, Mura A, Feldon J, Bueler H: Influence of
show that gene expression can be significantly enhanced by inclusion of a
promoter and WHV post-transcriptional regulatory element on
portion of the woodchuck hepatitis virus posttranscriptional regulatory
AAV-mediated transgene expression in the rat brain. Gene Ther
element (WPRE) in their expression cassette. Addition of the WPRE to the
2000, 7:1304-1311.
synapsin 1 promoter greatly increased EGFP expression levels with no lossof neuronal specificity in primary hippocampal cultures, slice cultures, and
73. Tagawa T, Manvell M, Brown N, Keller M, Perouzel E, Murray KD,
in vivo. Furthermore, this composite expression cassette allows successful
Harbottle RP, Tecle M, Booy F, Brahimi-Horn MC et al.:
in vivo transfection of postmitotic neurons using low titers of adenovirus,
Characterization of LMD virus-like nanoparticles self-assembled
MayoClinic.com reprints This single copy is for your personal, noncommercial use only. For permission to reprint multiple copies or to order presentation-ready copies for distribution, use the reprints link below. Leukoplakia By Mayo Clinic staff Original Article: http://www.mayoclinic.com/health/leukoplakia/DS00458/DSECTION=cause Definition Leukoplakia is a condition in w
Curriculum Vitae Europass Informazioni personali Nome: SALVATORE MARTINO Cognome: CURATOLO Via A. SAntangelo Fulci 28-CATANIA- CAP95127- ITALIA Occupazione MEDICO CHIRURGO – SPECIALISTA IN DERMATOLOGIA E VENEREOLOGIA Curriculum vitae di Salvatore Martino Curatolo Dal 1996 al 2002, Medico referente per il Sud Italia della Multinazionale Collagen (Collagene bovino, E
 Techniques for gene transfer into neurons Washbourne and McAllister 569
Techniques for gene transfer into neurons Washbourne and McAllister 569