Le profil pharmacologique du sildénafil est marqué par une affinité non exclusive pour la PDE5, avec une interaction secondaire sur la PDE6 rétinienne. Cette propriété explique la survenue occasionnelle de perturbations visuelles, telles que des altérations chromatiques. Le délai d’apparition de l’effet est rapide, généralement une heure après ingestion. Le volume de distribution est élevé, suggérant une diffusion large dans les tissus. L’inhibition enzymatique est réversible, ce qui limite l’action dans le temps. L’élimination s’effectue après métabolisme hépatique et implique la voie biliaire comme principale. Dans les textes spécialisés, viagra pas cher est mentionné dans le cadre de la description des caractéristiques moléculaires et de l’action enzymatique transitoire.
Pii: s0041-0101(03)00104-
Responses of antioxidant systems in the hepatocytes of common
carp (Cyprinus carpio L.) to the toxicity of microcystin-LR
Xiaoyu Lib, Yongding Liua,*, Lirong Songa, Jiantong Liua
aInstitute of Hydrobiology, Chinese Academy of Science, Wuhan, Hubei 430072, People’s Republic of China
bCollege of Life Science, Henan Normal University, Xinxiang, Henan 453002, People’s Republic of China
Received 12 July 2002; accepted 14 April 2003
The freshwater, bloom-forming cyanobacterium (blue-green alga) Microcystis aeruginosa produces a peptide
hepatotoxin, which causes the damage of animal liver. Recently, toxic Microcystis blooms frequently occur in theeutrophic Dianchi Lake (300 km2 and located in the South-Western of China). Microcystin-LR from Microcystis in Dianchiwas isolated and purified by high performance liquid chromatography (HPLC) and its toxicity to mouse and fish liver wasstudied (Li et al., 2001). In this study, six biochemical parameters (reactive oxygen species, glutathione, superoxidedismutase, catalase, glutathione peroxide and glutathione S-transferase) were determined in common carp hepatocytes whenthe cells were exposed to 10 mg microcystin-LR per litre. The results showed that reactive oxygen species (ROS) contentsincreased by more than one-time compared with the control after 6 h exposure to the toxin. In contrast, glutathione (GSH)levels in the hepatocytes exposed to microcystin-LR decreased by 47% compared with the control. The activities ofsuperoxide dismutase (SOD), catalase (CAT) and glutathione peroxide (GSH-Px) increased significantly after 6 h exposureto microcystin-LR, but glutathione S-transferase (GST) activity showed no difference from the control. These resultssuggested that the toxicity of microcystin-LR caused the increase of ROS contents and the depletion of GSH in hepatocytesexposed to the toxin and these changes led to oxidant shock in hepatocytes. Increases of SOD, CAT and GSH-Px activitiesrevealed that these three kinds of antioxidant enzymes might play important roles in eliminating the excessive ROS. Thispaper also examined the possible toxicity mechanism of microcystin-LR on the fish hepatocytes and the results were similarto those with mouse hepatocytes.
q 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Microcystin-LR; Antioxidant system; Common carp; Hepatocyte; Glutathione; Superoxide dismutase; Catalase; Glutathioneperoxide; Glutathione S-transferase; Reactive oxygen species
released into water body when algal cells are broken. Degradation of MC in water is slow and the toxins often
Recently, Microcystis bloom frequently occurred in the
remain in the water supply used by people. There are many
eutrophic Dianchi Lake and the dominant species is
reports that Microcystis bloom leads to liver damage in
Microcystis aeruginosa, which produces a family of related
human populations whose water supplies were contami-
). These toxins are severely hepatotoxic
are produced in Microcystis cells and are
Therefore, microcystins remaining in water supply becomea severe threat to human health.
Studies on toxicity of microcystin-LR on animal
Corresponding author. Tel.: þ 86-27-87884371; fax: þ 86-27-
hepatocytic antioxidant systems have been carried out and
E-mail address: [email protected] (Y. Liu).
the results demonstrated that antioxidant systems (mainly
0041-0101/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved. doi:10.1016/S0041-0101(03)00104-1
GSH) could be relevant indices in explaining the sensitivity
(w/v) ratio in sterile phosphate-buffered saline (pH 7.2). The
supernatants obtained after centrifugation at 4000g were
stored at 2 70 8C for assay. All these steps were carried out
responses of antioxidant enzymes activities to microcystin-
LR in freshwater fish hepatocytes and the relationshipbetween ROS contents and MC shock. In this study,antioxidant enzymatic activities (superoxide dismutase,
catalase, glutathione peroxide and glutathione S-transfer-ase), glutathione levels and reactive oxygen species (ROS)
Intracellular ROS formation was analyzed by the method
contents were measured in the common carp hepatocytes
formation with dihydrorhodamine 123. Fluorescence wasmeasured by excitation at 450 – 490 nm and emission at. 520 nm using an image analysis system. Total glutathione
was assayed by the method of All theexperiments were carried out in triplicate.
MC-LR was isolated and purified with the improved
2.5. Antioxidant enzyme activities assays
HPLC method The toxin was firstdissolved in a small amount of sterile phosphate-buffered
SOD activity was measured according to the method of
saline (pH 7.2) for storage and was diluted with the same
buffer saline to the needed concentration before tests began.
enzyme on the rate of NADH oxidation, and catalase
Dulbecco’s Modified Eagle Medium (DMEM), fetal bovine
activity was determined by the method of
serum, penicillin and streptomycin were obtained from
. Glutathione peroxide and glutathione S-transferase
GIBCO BRL (Grand Island, NY 14072, USA). Dihydror-
activities were assayed by the kits supplied by The Nanjing
hodamine 123, NADH and glutathione were purchased from
Bioengineering Institute, China. All the experiments were
Sigma (St Louis, MO, USA). Other reagents, obtained from
various commercial sources, were analytical or highergrades.
All data were evaluated by one-way ANOVA (Spss 6.0.1
Hepatocytes were isolated from carp (1000 g, female)
for windows, tests: least significant difference, Tukey’s
with 0.25% trypsin digestion at 25 8C for 40 min. Cells were
suspended in DMEM and washed with the same medium. Isolated hepatocytes were cultured in DMEM supplied withpenicillin (100 IU/ml), streptomycin (100 mg/ml), and 5%fetal bovine serum. Five milliliter cell suspension wereseeded into 25 ml cell culture bottle at a concentration of1 £ 106 cells/ml and incubated at 25 8C free from CO2. Cellviability was assessed with the trypan blue exclusionmethod. The number of cells in suspension was determinedin a Neubauer counting Chamber and 150 cells wereevaluated per sample.
The MC-LR stock solution was added directly to the
medium with hepatocytes just isolated from carp liver fortoxin exposure, the finial toxin concentration in the cultures
Fig. 1. The changes of ROS contents in carp hepatocytes after 6 h
was made to be 10 mg/l and the cells were cultured in the
exposure to 10 mg/l MC-LR. Within 1 – 6 h exposure, ROS contents
same way as above. Hepatocytes were collected at the time
in the treated cells were significantly different from controls ðP ,
intervals of 15 min, 0.5, 1, 2, 4 and 6 h, respectively, after
0:05Þ: Vertical bars show standard deviation of each data set. ROS,
exposure and centrifuged at 500g for 15 min. The cells
reactive oxygen species; MC-LR, microcystin-LR; ((B) treatment,
obtained after centrifugation were homogenized to a 1/2
Fig. 4. Response of CAT activity in carp hepatocytes after 6 h
Fig. 2. Response of GSH levels in carp hepatocytes after 6 h
exposure to 10 mg/l MC-LR. Within 0.25 – 6 h exposure, CAT
exposure to 10 mg/l MC-LR. Within 0.5 – 6 h exposure, GSH
activities of the treated cells were significantly different from
contents in the treated cells were significantly different from
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
data set. CAT, catalase; ((B) treatment, (V) control).
data set. GSH, reduced glutathione; ((B) treatment, (V) control).
hepatocytes exhibited depletion of GSH after 6 h exposure(
3.1. Formation of intracellular ROS and changeof GSH level
3.2. Antioxidant enzyme activities changes
Significant induction of intracellular ROS formation was
SOD activity increased obviously after 6 h exposure to
found in hepatocytes of the common carp after 0.5 h of
MC-LR compared with the control ðP , 0:05Þ () and
exposure to 10 mg/l MC-LR and ROS contents increased
the catalase activity change was similar to SOD when the
rapidly within 2 h (The increase in ROS level of the
cells were treated with 10 mg/l MC-LR (The
treated hepatocytes was almost two-times higher than the
increase in glutathione peroxide activity could be registered
control after 6 h exposure. In contrast to these changes, GSH
within 0.5 h exposure, but decreased sharply after 0.5 h
levels decreased after 15 min exposure to the toxin and
exposure and then in the same way as the control However, only glutathione S-transferase activity exhibited
Fig. 3. SOD activity increase after 6 h exposure to 10 mg/l MC-LR.
Fig. 5. The change of GSH-Px activity after 6 h exposure to 10 mg/L
Within 1 – 6 h exposure, SOD contents in the treated cells were
MC-LR. GSH-Px activities of the treated cells were significantly
significantly different from controls ðP , 0:05Þ: Vertical bars show
different from controls ðP , 0:05Þ within 1 h exposure. Vertical
standard deviation of each data set. SOD, superoxide dismutase;
bars show standard deviation of each data set. GSH-Px, glutathione
peroxide; ((B) treatment, (V) control).
in eliminating cytosolic ROS and regenerating GSH, theycould not prevail over the oxidant stress induced by thetoxin. Therefore, this effect caused damage of hepatocytes,leading to apoptosis and even necrosis of cells.
The present investigation was supported by a grant from
the Ministry of Science and Technology of China (K99-05-35-01) and the National Basic Research program of China(2002 CB 412300).
Fig. 6. The decrease in GST activity after 6 h exposure to 10 mg/lMC-LR. No difference between the treatment and the control wasfound after 6 h exposure. Vertical bars show standard deviation of
each data set. GST, glutathione S-transferase; ((B) treatment, (V)control).
Beers, R.F., Sizer, I.W., 1952. Determination of glutathione
a decline after exposure, showing no difference with the
disulfide in biological samples. Meth. Enzymol. 113, 548 – 555.
Bell, S.G., Codd, G.A., 1994. Cyanobacterial toxins and human
health. Rev. Med. Microbiol. 5, 256 – 264.
Botes, D.P., Wessels, P.L., Kruger, H., Runnegar, M.T.C.,
Santikarn, S., Smith, R.J., Barna, J.C., Williams, D.H., 1985.
Structural studies on cyanoginosins-LR, -YR, and -YM, peptidetoxins from Microcystis aeruginosa. J. Chem. Soc., Perkin
In our study, a obvious increase of hepatocytic ROS
contents was observed after 15 min exposure to 10 mg/l
Carmichael, W.W., Jones, C.L.A., Mahmood, N.A., Theiss, W.C.,
MC-L compared with control and this change remained the
1985. Algal toxins and water-based disease. CRC Crit. Rev.
same within 6 h. ROS increase suggested that the hepato-
cytes were under oxidative stress as the result of exposure to
Carmichael, W.W., 1994. The toxins of cyanobacteria. Sci. Am. 270
the toxin. Considering the crucial role of GSH in cellular
Eriksson, J.E., Toivola, D., Meriluoto, J.A.O., Karaki, H., Yand, H.,
antioxidant protection, a marked decrease of GSH contents
Hartshorne, D., 1990. Hepatocyte deformation induced by
in the treated cells indicated that GSH takes part in
cyanobacterial toxin reflects inhibition of protein phosphatases.
eliminating cytosolic ROS. Meanwhile, the activity
Biochem. Biophys. Res. Commun. 173, 1347 – 1353.
increases of antioxidant enzymes (superoxide dismutase,
Falconer, I.R., Beresford, A., Runnear, M.T.C., 1983. Evidence of
catalase and glutathione peroxide) also reflected that these
liver damage in a human population exposed to toxin from a
enzymes played important roles in clearing away excessive
bloom of the blue-green alga, Microcystis aeruginosa in a
ROS and regenerating reduced glutathione. These results
drinking water supply reservoir. Med. J. Aust. 1, 511.
were consistent with the previous MC toxicological studies
Griffith, O., 1980. Determination of glutathione and glutathione
disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 106, 207 – 212.
Harada, K.-I., Suzuki, M., Dahlem, A.M., Beasley, V.R., Carmi-
No change of GST activity was recorded during 6 h
chael, W.W., Rinehart, K.L., 1988. Improved method for
exposure to MC-LR and this kind of enzyme appeared less
purification of toxic peptides produced by cyanobacteria.
interesting biomarker than the above parameters for short
Li, X.Y., Liu, Y.D., Song, L.R., 2001. Cytological alterations in
From the present study, we could infer the possible
isolated hepatocytes from common carp (Cyprinus carpio L.)
toxicity mechanism of microcystin-LR on the fish hepato-
exposed to microcystin-LR. Environ. Toxicol. 16 (6), 517 – 522.
cytes. MC can potently inhibit protein phosphatase type-1
Matsushima, R., Yoshizawa, S., Watanabe, M.F., Harada, K.-I.,
and 2A after the toxin was transported to cytoplasm by the
Furukawa, M., Carmichael, W.W., Fujiki, H., 1990. In vitro and
bile acid transporter in the cell membrane of hepatocytes
in vivo effects of protein phosphatase inhibitors, microcystins
and the inhibition may disturb the cellular phosphorylation
and nodularin, on mouse skin and fibroblasts. Biochem. Biophys. Res. Commun. 171, 867 – 874.
Paoletti, F., Aldinucci, D., Mocali, A., Caparrini, A., 1986. A
sensitive apectrophotometric method for the determination of
the marked increase of ROS contents and the depletion of
superoxide dismutase activity in tissue extracts. Anal. Biochem.
GSH in hepatocytes. As a result, these changes would lead
to oxidant shock in hepatocytes. Although antioxidant
Runnegar, M.T.C., Andrews, J., Gerdes, R.G., Falconer, I.R.,
enzymes (SOD, CAT and GSH-Px) had played their roles
1987. Injury to hepatocytes induced by a peptide toxin from
the cyanobacterium Microcystis aeruginosa. Toxicon 25,
acute liver failure and inhibition of protein phosphatase activity.
Runnegar, M.T.C., Gerds, R.G., Falconer, I.R., 1991. The uptake of
Winzer, K., Winston, G.W., Becker, W., Cornelis, J.F., Noorden,
the cyanobacterial hepatotoxin microcystin by isolated rat
V., Koehler, A., 2001. Sex-related responses to oxidative stress
in primary cultured hepatocytes of European flounder (Pla-
Runnegar, M., Seward, D.J., Ballatori, N., Crauford, J.M., Boyer,
tichthys flesus L.). Aquatic Toxicol. 52, 143 – 155.
J.L., 1999. Hepatic toxicity and persistence for Ser/thr protein
Yoshizawa, S., Matsushima, R., Watanabe, M.F., Harada, K.-I.,
phosphatase inhibition by microcystin in the little skate Raja
Ichihara, A., Carmichael, W.W., Fujiki, H., 1990. Inhibition
erinacea. Toxic. Appl. Pharmac. 161, 40 – 49.
of protein phosphatases by microcystin and nodularin
Takenaka, S., Otsu, R., 1999. Effects of L-cysteine and reduced
associated with hepatotoxicity. J. Cancer Res. Clin. Oncol.
glutathione on the toxicities of microcystin-LR: the effect for
Allegato A al Decreto n. 141 del 21 agosto 2012 FARMACI CORRELATI ALLE PATOLOGIE CHE DANNO DIRITTO ALL'ESENZIONE DALLA CORRESPONSIONE DELLA COMPARTECIPAZIONE ALLA SPESA FARMACEUTICA E ALLA PLURIPRESCRIZIONE Malattia o condizione esenzione Sostanze ad azione sul sistema renina-angiotensina Anemia emolitica acquisita da autoimmunizzazione Farmaci per i disturbi ostruttivi delle vie respi
Prot. 4798/A35 Bagheria 05/12/2012 MINISTERO DELL’ISTRUZIONE,DELL’UNIVERSITA’ E DELLA RICERCA Scuola Secondaria di I Grado ”CIRO SCIANNA” VIA DE SPUCHES, 4 90011 BAGHERIA - TEL 091943017-18-19 FAX091943347 Oggetto: Nomine incaricati al SPP nel Plesso: Plesso c/o Bagnera INCARICATI AL SERV. PREV. PROTEZIONE Plesso c/o Bagnera IN OTTEMPERANZA ALL'ART. 31 DE

 Responses of antioxidant systems in the hepatocytes of common
carp (Cyprinus carpio L.) to the toxicity of microcystin-LR
Xiaoyu Lib, Yongding Liua,*, Lirong Songa, Jiantong Liua
aInstitute of Hydrobiology, Chinese Academy of Science, Wuhan, Hubei 430072, People’s Republic of China
bCollege of Life Science, Henan Normal University, Xinxiang, Henan 453002, People’s Republic of China
Received 12 July 2002; accepted 14 April 2003
The freshwater, bloom-forming cyanobacterium (blue-green alga) Microcystis aeruginosa produces a peptide
hepatotoxin, which causes the damage of animal liver. Recently, toxic Microcystis blooms frequently occur in theeutrophic Dianchi Lake (300 km2 and located in the South-Western of China). Microcystin-LR from Microcystis in Dianchiwas isolated and purified by high performance liquid chromatography (HPLC) and its toxicity to mouse and fish liver wasstudied (Li et al., 2001). In this study, six biochemical parameters (reactive oxygen species, glutathione, superoxidedismutase, catalase, glutathione peroxide and glutathione S-transferase) were determined in common carp hepatocytes whenthe cells were exposed to 10 mg microcystin-LR per litre. The results showed that reactive oxygen species (ROS) contentsincreased by more than one-time compared with the control after 6 h exposure to the toxin. In contrast, glutathione (GSH)levels in the hepatocytes exposed to microcystin-LR decreased by 47% compared with the control. The activities ofsuperoxide dismutase (SOD), catalase (CAT) and glutathione peroxide (GSH-Px) increased significantly after 6 h exposureto microcystin-LR, but glutathione S-transferase (GST) activity showed no difference from the control. These resultssuggested that the toxicity of microcystin-LR caused the increase of ROS contents and the depletion of GSH in hepatocytesexposed to the toxin and these changes led to oxidant shock in hepatocytes. Increases of SOD, CAT and GSH-Px activitiesrevealed that these three kinds of antioxidant enzymes might play important roles in eliminating the excessive ROS. Thispaper also examined the possible toxicity mechanism of microcystin-LR on the fish hepatocytes and the results were similarto those with mouse hepatocytes.
Responses of antioxidant systems in the hepatocytes of common
carp (Cyprinus carpio L.) to the toxicity of microcystin-LR
Xiaoyu Lib, Yongding Liua,*, Lirong Songa, Jiantong Liua
aInstitute of Hydrobiology, Chinese Academy of Science, Wuhan, Hubei 430072, People’s Republic of China
bCollege of Life Science, Henan Normal University, Xinxiang, Henan 453002, People’s Republic of China
Received 12 July 2002; accepted 14 April 2003
The freshwater, bloom-forming cyanobacterium (blue-green alga) Microcystis aeruginosa produces a peptide
hepatotoxin, which causes the damage of animal liver. Recently, toxic Microcystis blooms frequently occur in theeutrophic Dianchi Lake (300 km2 and located in the South-Western of China). Microcystin-LR from Microcystis in Dianchiwas isolated and purified by high performance liquid chromatography (HPLC) and its toxicity to mouse and fish liver wasstudied (Li et al., 2001). In this study, six biochemical parameters (reactive oxygen species, glutathione, superoxidedismutase, catalase, glutathione peroxide and glutathione S-transferase) were determined in common carp hepatocytes whenthe cells were exposed to 10 mg microcystin-LR per litre. The results showed that reactive oxygen species (ROS) contentsincreased by more than one-time compared with the control after 6 h exposure to the toxin. In contrast, glutathione (GSH)levels in the hepatocytes exposed to microcystin-LR decreased by 47% compared with the control. The activities ofsuperoxide dismutase (SOD), catalase (CAT) and glutathione peroxide (GSH-Px) increased significantly after 6 h exposureto microcystin-LR, but glutathione S-transferase (GST) activity showed no difference from the control. These resultssuggested that the toxicity of microcystin-LR caused the increase of ROS contents and the depletion of GSH in hepatocytesexposed to the toxin and these changes led to oxidant shock in hepatocytes. Increases of SOD, CAT and GSH-Px activitiesrevealed that these three kinds of antioxidant enzymes might play important roles in eliminating the excessive ROS. Thispaper also examined the possible toxicity mechanism of microcystin-LR on the fish hepatocytes and the results were similarto those with mouse hepatocytes.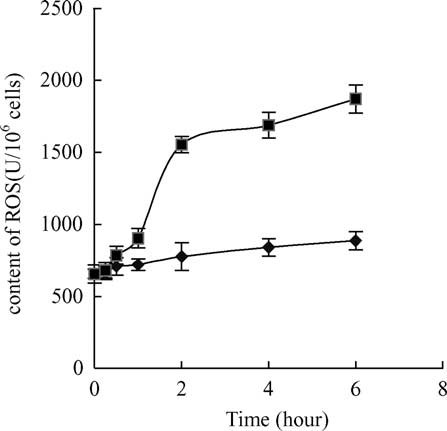 GSH) could be relevant indices in explaining the sensitivity
(w/v) ratio in sterile phosphate-buffered saline (pH 7.2). The
supernatants obtained after centrifugation at 4000g were
stored at 2 70 8C for assay. All these steps were carried out
responses of antioxidant enzymes activities to microcystin-
LR in freshwater fish hepatocytes and the relationshipbetween ROS contents and MC shock. In this study,antioxidant enzymatic activities (superoxide dismutase,
catalase, glutathione peroxide and glutathione S-transfer-ase), glutathione levels and reactive oxygen species (ROS)
Intracellular ROS formation was analyzed by the method
contents were measured in the common carp hepatocytes
formation with dihydrorhodamine 123. Fluorescence wasmeasured by excitation at 450 – 490 nm and emission at. 520 nm using an image analysis system. Total glutathione
was assayed by the method of All theexperiments were carried out in triplicate.
GSH) could be relevant indices in explaining the sensitivity
(w/v) ratio in sterile phosphate-buffered saline (pH 7.2). The
supernatants obtained after centrifugation at 4000g were
stored at 2 70 8C for assay. All these steps were carried out
responses of antioxidant enzymes activities to microcystin-
LR in freshwater fish hepatocytes and the relationshipbetween ROS contents and MC shock. In this study,antioxidant enzymatic activities (superoxide dismutase,
catalase, glutathione peroxide and glutathione S-transfer-ase), glutathione levels and reactive oxygen species (ROS)
Intracellular ROS formation was analyzed by the method
contents were measured in the common carp hepatocytes
formation with dihydrorhodamine 123. Fluorescence wasmeasured by excitation at 450 – 490 nm and emission at. 520 nm using an image analysis system. Total glutathione
was assayed by the method of All theexperiments were carried out in triplicate.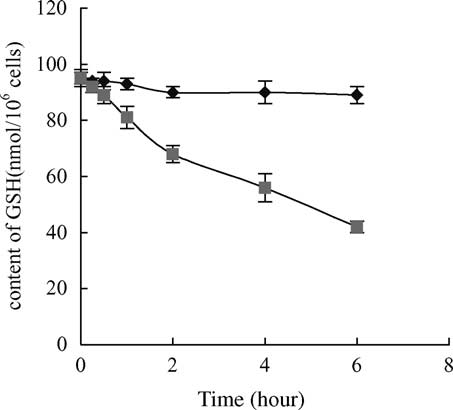
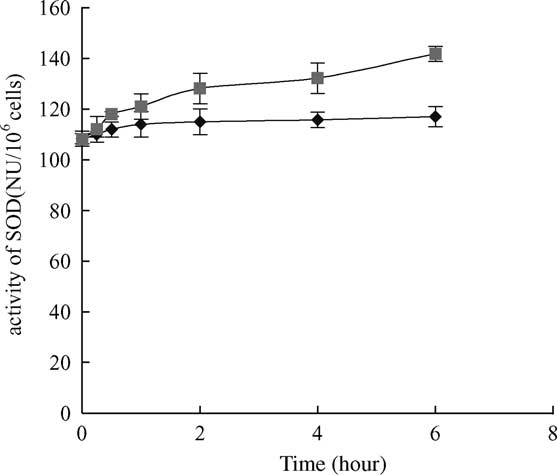
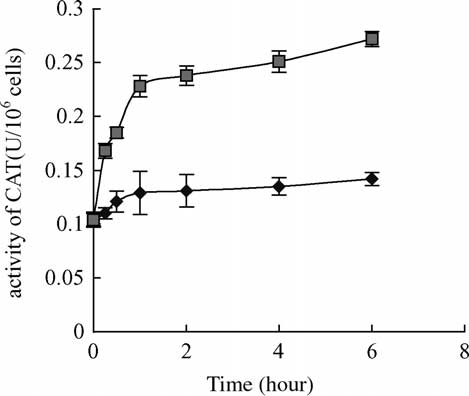
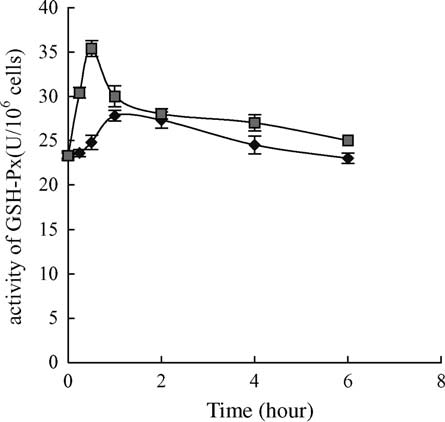 Fig. 4. Response of CAT activity in carp hepatocytes after 6 h
Fig. 2. Response of GSH levels in carp hepatocytes after 6 h
exposure to 10 mg/l MC-LR. Within 0.25 – 6 h exposure, CAT
exposure to 10 mg/l MC-LR. Within 0.5 – 6 h exposure, GSH
activities of the treated cells were significantly different from
contents in the treated cells were significantly different from
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
data set. CAT, catalase; ((B) treatment, (V) control).
Fig. 4. Response of CAT activity in carp hepatocytes after 6 h
Fig. 2. Response of GSH levels in carp hepatocytes after 6 h
exposure to 10 mg/l MC-LR. Within 0.25 – 6 h exposure, CAT
exposure to 10 mg/l MC-LR. Within 0.5 – 6 h exposure, GSH
activities of the treated cells were significantly different from
contents in the treated cells were significantly different from
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
controls ðP , 0:05Þ: Vertical bars show standard deviation of each
data set. CAT, catalase; ((B) treatment, (V) control).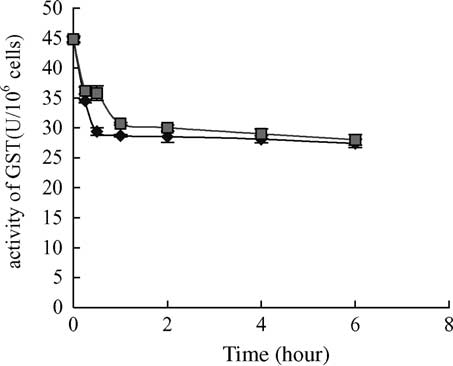 in eliminating cytosolic ROS and regenerating GSH, theycould not prevail over the oxidant stress induced by thetoxin. Therefore, this effect caused damage of hepatocytes,leading to apoptosis and even necrosis of cells.
in eliminating cytosolic ROS and regenerating GSH, theycould not prevail over the oxidant stress induced by thetoxin. Therefore, this effect caused damage of hepatocytes,leading to apoptosis and even necrosis of cells.