Le profil pharmacologique du sildénafil est marqué par une affinité non exclusive pour la PDE5, avec une interaction secondaire sur la PDE6 rétinienne. Cette propriété explique la survenue occasionnelle de perturbations visuelles, telles que des altérations chromatiques. Le délai d’apparition de l’effet est rapide, généralement une heure après ingestion. Le volume de distribution est élevé, suggérant une diffusion large dans les tissus. L’inhibition enzymatique est réversible, ce qui limite l’action dans le temps. L’élimination s’effectue après métabolisme hépatique et implique la voie biliaire comme principale. Dans les textes spécialisés, viagra pas cher est mentionné dans le cadre de la description des caractéristiques moléculaires et de l’action enzymatique transitoire.
Pii: s0014-5793(01)03050-2
Cadmium inhibits epoxidation of diatoxanthin to diadinoxanthin in the
xanthophyll cycle of the marine diatom Phaeodactylum tricornutum
Martine BertrandaYbY*, Beno|ªt SchoefsbY1, Pavel Si¡elc, Karel Rohacekc, Istvan Molnarc
aInstitut National des Sciences et Techniques de la Mer, Conservatoire National des Arts et Me¨tiers, BP 324, F-50103 Cherbourg Cedex, France
bLaboratory of Biomembranes, University of South Bohemia, Branisovska 31, CZ-370 05 Ceske Budejovice, Czech Republic
cLaboratory of Photosynthesis, Institute of Plant Molecular Biology, Academy of Sciences of the Czech Republic, Branisovska 31,
CZ-370 05 Ceske Budejovice, Czech Republic
Received 6 August 2001; accepted 4 October 2001
Abstract Cd has pleiotropic effects on plant physiology and in
higher plants and green algae, the xanthophyll cycle consists
particular on photosynthesis. It has not been established yet if Cd
in the reversible conversion of violaxanthin to zeaxanthin via
alters the functioning of the xanthophyll cycle. To answer this
antheraxanthin, whereas in chromophytes, it consists in the
question, an exponentially growing culture of the marine diatom
interconversion of diadinoxanthin (DD) and diatoxanthin
Phaeodactylum tricornutum was incubated with Cd (20 mg/l) for
(DT) ([9], reviewed in [8]). The activity of this cycle has
24 h and irradiated with a light activating the xanthophyll cycle,
been correlated with the thermal dissipation of the excess en-
which in diatoms, consists of the reversible deepoxidation of
ergy at the antenna side of the photosynthetic apparatus
diadinoxanthin to diatoxanthin. The measurements show that the
deepoxidation step is not influenced by Cd. In contrast, the Cd
In this contribution we report evidence, based on absor-
concentration used sharply inhibits the epoxidation of diatox-
bance and £uorescence measurements and pigment quanti¢-
anthin to diadinoxanthin. ß 2001 Federation of European Bio-
chemical Societies. Published by Elsevier Science B.V. All
cations, for the partial inhibition by Cd of the epoxidation
step of the xanthophyll cycle in the marine diatom Phaeodac-
Key words: Algae; Microalgae; Cd; Heavy metal stress;
The experiments were performed with the marine diatom P. tricor-
nutum BOHLIN COUGHLAN/-632 axenic strain from the collection
at the Institute of Botany at Trebon, Czech Republic. The cells were
There is considerable concern about the increasing levels of
grown in a medium (pH 8.3) containing ¢ltered reconstituted sea
heavy metals in the environment and the potential harmful
water (33 g/l of commercial salts from the Red Sea), Na2HPO4 (140
WM), NaNO3 (1560 WM), Na2SiO3 (130 WM), CuSO4 (1.4 WM), Co-
e¡ects of these elements on living organisms. This is especially
(NO3)2W6H2O (1.6 WM), MnCl2 (3.2 WM), Mo (0.7 WM), ZnCl2 (0.3
important in the case of heavy metals like Cd, which have no
WM), Fe^EDTA (100 WM), vitamin B1 (54 WM), vitamin B12 (0.7 WM)
and vitamin H (700 WM). The chemicals (P.A. quality) were purchased
The photosynthetic process is very sensitive to Cd: it re-
by Fluka, and the water was of Milli-Q quality (Millipore).
Erlenmeyer £asks containing 100 ml of culture medium were inoc-
duces growth, inhibits chlorophyll (Chl) biosynthesis, de-
ulated with 25 ml of a mother diatom culture. The microalgae were
presses the photosynthetic carbon assimilation, etc. ([2^5], re-
grown under a light^dark cycle (16 h/8 h, illumination: £uorescent
viewed in [6]). To our knowledge, only scant information is
tubes, 40 Wmol photons/m2/s PAR) in a phytotron thermostated at
available about the in£uence of Cd on the xanthophyll cycle,
an important photoprotection process of photosynthesis trig-
Di¡erent concentrations of CdCl2 (0^25 mg/l of Cd) were applied
the 4th day after inoculation (exponential phase). Measurements were
gered under high-light conditions ([7], reviewed in [8]). In
2.2. Induction of the xanthophyll cycle and £uorescence measurements
*Corresponding author. Fax: (33)-2-33 88 73 39.
Chl £uorescence induction kinetics were measured using a PAM
E-mail address: [email protected] (M. Bertrand).
101-103 £uorimeter (Walz, Germany). The weak modulated measur-
ing light (ML; 1.6 kHz) was delivered by a weak blue LED (V = 450
1 Present address: Plasticite¨ et Expression des Ge¨nomes Microbiens,
nm, Nichia, Japan). The light was ¢ltrated through a Corning 4-96
Universite¨ Joseph Fourier, CNRS-FRE 2383, BP 53, F-38041
¢lter (380 nm 6 V 6 600 nm). The detector was protected by a red
Before irradiation of a cell suspension by actinic light (AL), each
Abbreviations: AL, actinic light; Chl, chlorophyll; DD, diadinoxan-
sample was dark-adapted for 15 min and irradiated by a weak far-red
thin; DT, diatoxanthin; DTT, dithiothreitol; F0, minimum chloro-
(FR) light (V = 735 nm, V10 Wmol/m2/s) for 5 s to re-oxidize the
phyll £uorescence level; F0P, minimum chlorophyll £uorescence level
electron transport carriers. Then, the minimum Chl £uorescence level
of the light-adapted state; FM, maximum chlorophyll £uorescence
(F0 ) corresponding to open photosystem II (PSII) reaction centers
level; FMP, maximum chlorophyll £uorescence level of the light-
was recorded using ML. Next, one polychromatic saturation pulse
adapted state; Fv, variable £uorescence; FR, far-red; ML, modulated
(SP; 1000 Wmol/m2/s, 1 s, light source: KL 1500 lamp, Walz, Ger-
measuring light; NPQ, non-photochemical quenching; PSII, photo-
many) was applied to determine the maximum £uorescence level (FM).
After three min of relaxation in the dark, the suspension was illumi-
0014-5793 / 01 / $20.00 ß 2001 Federation of European Biochemical Societies. Published by Elsevier Science B.V. All rights reserved.
PII: S 0 0 1 4 - 5 7 9 3 ( 0 1 ) 0 3 0 5 0 - 2
M. Bertrand et al./FEBS Letters 508 (2001) 153^156
nated with AL (1000 Wmol/m2/s, light source: KL 1500 lamp, Walz,
Germany) for 7 min to induce £uorescence quenching. The intensity
of AL was high enough to keep the PSII closed throughout the ex-
posure and thus the observed £uorescence decline only represents
When AL was switched o¡ and FR was applied, the minimum Chl
£uorescence level corresponding to the light adapted state (F0P) was
determined. Then, dark relaxation of both F0P and maximum Chl
£uorescence level of the light-adapted state (FMP) were measured.
During this period, light pulses were given at a regular interval of
5 min. The ¢rst one was triggered 1 min after switching o¡ the AL
(see Fig. 1). Variable £uorescence (Fv) was calculated as (FM3F0).
2.3. Pigment extraction and separation by HPLC
One milliliter of cells was taken at selected moments of the light^
dark treatment described above (see Fig. 2). The cells suspension was
spun down at 4000Ug. The supernatant was discarded and the pig-
ments were extracted with 800 Wl tetrahydrofuran. The extract was
centrifuged at 15 000Ug and the pellet was extracted again with 400 Wl
acetone. The operation was repeated once, but using 800 Wl methanol.
After the last centrifugation, the pellet was colorless. The colored
supernatants were pooled, ¢ltered on a PTFE ¢lter membrane (pore
Fig. 2. Variations of the relative amount of DT during the light^
diameter: 0.45 Wm), and dried under a nitrogen stream. The dried
dark sequence applied on P. tricornutum grown in the absence or
pigments were either directly used for HPLC analysis or stored at
380³C. All the operations were performed in a cold room and under
a dim green light to avoid pigment alterations [12].
The dried samples were dissolved in 250 Wl of methanol and ana-
periments using dithiothreitol (DTT), the algae were incubated in the
lyzed according to [13]. The diode-array detector, the pump system
presence of the compound (1 mM) in the dark for 20 min.
and the column were described elsewhere [14]. All solvents were of
HPLC quality. DD and DT were detected on the basis of the chro-
matograms recorded at 445 and 451 nm, respectively.
3.1. In£uence of Cd concentration on the cell growth
Light-induced absorbance di¡erence spectra were measured on cell
The increase of Cd concentration in the culture medium
suspensions with the laboratory-built £ash spectrophotometer com-
reduced the growth rate: the higher the concentration in
posed of a microsecond Xe £ash lamp (FX-1160, EG4G, USA, £ash
duration of 1 Ws), an imaging monochromator (MS257, Oriel, USA)
Cd, the lower the rate (data not shown). The growth rate
and an acquisition and triggering unit FL100 (Photon System Instru-
was reduced by 50% using a concentration of 20 mg Cd/l. A
ments, Czech Republic). The detector was composed of two large area
similar value was reported in [15]. Therefore, this concentra-
photodiode arrays (S4111, Hamamatsu) for sample and reference.
tion was used for all the experiments.
Measuring light pulse was ¢ltered using a combination of GG475
¢lter (Schott, V s 480 nm) and a Corning 4-96 ¢lter (380
nm 6 V 6 600 nm), divided to sample and reference channels by a
3.2. Cd slows down the £uorescence yield relaxation
broadband beam splitter and focused to sample and reference cells.
In photosynthetic organisms, the relative decrease of the
After passage of light through a sample and a reference, both light
£uorescence yield is correlated with the activity of the xantho-
beams were focused to two bundles of optical ¢bers and directed to
phyll cycle (higher plants: [16], diatoms: [17,18]) although it
the monochromator input. This experimental setup gave a noise as
low as 2U1034 per one £ash with a spectral resolution of 2.1 nm per
cannot be fully explained by the xanthophyll conversion
one spectral point. Each absorbance measurement represents an ac-
[10,17,18]. Fig. 1 presents typical Chl £uorescence induction
cumulation of 10 spectra measured within 1 s. The Chl concentration
kinetics of non-treated and Cd-treated cells. The comparison
was approximately 20 mg/l. To reach this concentration, the cells were
of the ¢gures presented in Table 1 clearly indicated that Cd
sedimented without centrifugation (15 min in darkness). For the ex-
has no e¡ect on the maximum yield of PSII photochemistry
(Fv/FM) or the NPQ mechanism ((FM3FMP)/FMP), which did
not di¡er signi¢cantly in Cd-treated algae. However, the rate
of the F0P relaxation in the dark is reduced by 58% in the Cd-
It has been shown that the F0 quenching is of non-photo-
chemical nature. It could be induced by the formation of high-
energy state of thylakoid membranes [19,20], photoinhibition
of PSII reaction centers and/or by the xanthophyll cycle ac-
tivity [21,22]. Depression of £uorescence intensity by photo-
inhibition is accompanied by a pronounced decrease of the
Fv/FM ratio, which is not observed in the presence of Cd
(Table 1). High-energy state of thylakoid membranes of high-
E¡ects of the Cd addition on the Chl £uorescence induction param-
Fig. 1. Typical Chl £uorescence induction kinetics recorded with liv-
ing cells of P. tricornutum in the absence or presence of Cd (20 mg/
M. Bertrand et al./FEBS Letters 508 (2001) 153^156
er plants, and related £uorescence decrease, usually relax
within 1 min after switching o¡ AL. For Phaeodactylum cells,
the relaxation lasted 10^30 min (Fig. 1). This duration corre-
sponds to the rate of DT epoxidation [23]. Hence, the decrease
in the rate of F0 relaxation, observed in the presence of Cd,
may re£ect an inhibition of the xanthophyll epoxidation re-
3.3. Inhibition of the epoxidation step of the xanthophyll cycle
In order to determine whether the slowing down of the F0
relaxation rate in Cd-treated cells is correlated with a change
in the xanthophyll cycle activity, the previous experiment was
repeated and the variations of the DD and DT amounts were
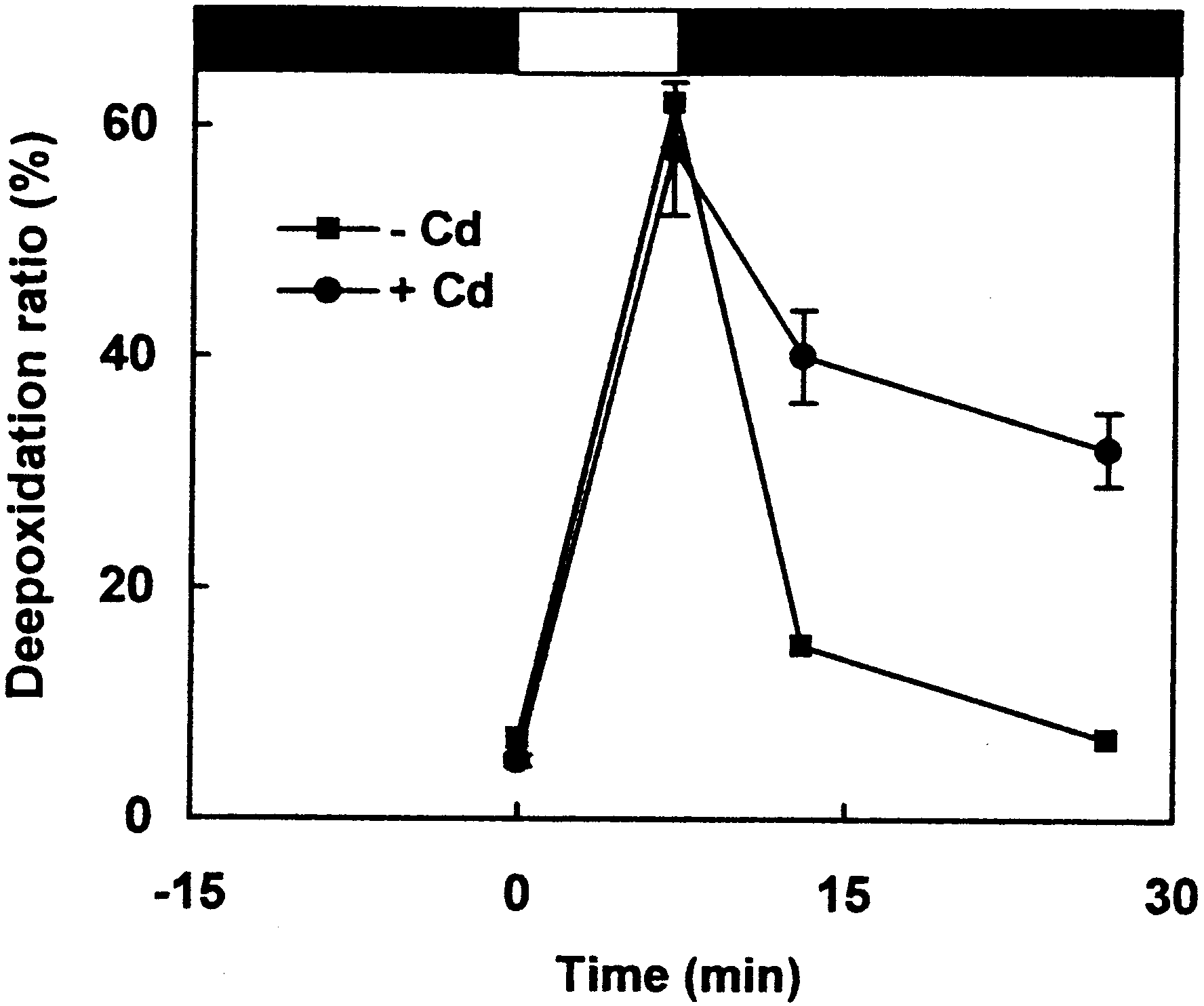
measured after HPLC separation. Fig. 2 presents the varia-
tions of the relative amount of DT during the light^dark se-
quence in the absence or in the presence of Cd. Regardless of
the presence of Cd, the same level of deepoxidation was
reached at the end of the high-light illumination. In contrast,
the reversion of the xanthophyll cycle was strongly slowed
down in the presence of Cd. Remarkably, the inhibition of
epoxidation rate was very similar in magnitude (approxi-
mately 60%) to that of the relaxation rate (Table 1). This
de¢nitively con¢rms that both phenomenons are tightly
3.4. Measurements of the xanthophyll cycle in vivo
It has been shown with higher plants that the deepoxidation
of violaxanthin to zeaxanthin is re£ected by an increase of the
absorbance at approximately 505 nm [24^26]. In order to
Fig. 3. E¡ect of Cd on the absorbance di¡erence spectra of P. tri-
cornutum (the control stayed in darkness). A: (b) 3Cd+7 min AL;
verify whether the activity of the xanthophyll cycle in diatoms
(F) 3Cd+7 min AL+40 min darkness; (R) 3Cd+DTT+7 min AL.
is also correlated to similar absorbance changes, we recorded
B: (b) +Cd+7 min AL; (F) +Cd+7 min AL+7 min darkness; (R)
absorbance spectra at room temperature during the light^dark
sequence (see Figs. 1 and 2). The spectra were subtracted from
the spectrum recorded before illumination. The 7 min AL,
ment quanti¢cation data reveals that the rate of F0 relaxation
which triggers the conversion of DD to DT (Fig. 2), induced
re£ects the rate of DT epoxidation. Similarly to higher plants,
a large increase of the absorbance at approximately 465 and
the xanthophyll cycle in diatoms can be followed by absor-
500 nm (Fig. 3A, circles). The absorbance changes were fully
bance variations at approximately 500 nm.
reversible in the dark (Fig. 3A, squares). This result clearly
establishes that the activity of the xanthophyll cycle in dia-
Acknowledgements: The authors thank the Ministery of Education
toms can be followed by measuring absorbance modi¢cations
and the Grant Agency of the Czech Republic for ¢nancial support.
at 500 nm. To obtain an additional evidence for this conclu-
B.S. is a research associate at the CNRS (France).
sion, we incubated the algae with DTT, a speci¢c inhibitor of
violaxanthin deepoxidase in higher plants [24] and DD deep-
oxidase in diatoms [18]. In this condition, the absorbance
changes at approximately 500 nm were totally inhibited
[1] Lane, T.W. and Morel, F.M.M. (2000) Proc. Natl. Acad. Sci.
(Fig. 3B, triangles). This result suggests that the two deep-
[2] Bo«ddi, B., Oravecz, A.R. and Lehoczki, E. (1995) Photosynthe-
oxidases have a similar mode of functioning. This conclusion
is in line with the recent ¢nding of Jakob and collaborators
[3] Sheoran, I.S., Singhal, H.R. and Singh, R. (1990) Photosynth.
[27] who showed that DD deepoxidase can also use violaxan-
[4] Fargasova, A. (1999) Biologia 54, 303^308.
[5] Reinhold, D., Hofner, W. and Kohler, W. (1994) Z. P£anzener-
Fig. 3B presents the absorbance changes measured with
algae grown in the absence or presence of Cd. As expected,
[6] Bertrand, M., Guary, J.C. and Schoefs, B. (2001) in: Handbook
regardless of the presence or absence of Cd, the absorbance
of Plant and Crop Physiology (Pessarakli, M., Ed.), pp. 751^762,
changes at 500 nm were equivalent after the light treatment
(Fig. 3B, circles). In contrast, the decrease of the absorbance
[7] Demmig-Adams, B. (1990) Biochim. Biophys. Acta 1020, 1^24.
[8] Rmiki, N-E., Schoefs, B. and Lemoine Y. (1999) in: Handbook
at 500 nm, which re£ects the epoxidation of DT to DD, was
of Plant and Crop Stress (Pessarakli, M., Ed.), pp. 465^482,
slower in the presence of Cd (Fig. 3B, compare squares (+Cd)
[9] Stansky, H. and Hager, A. (1970) Arch. Mikrobiol. 73, 315^323.
3.4.1. To summarize. Cd does not in£uence the deepoxi-
[10] Olaizola, M., La Roche, J., Kolber, Z. and Falkowski, P.G.
(1994) Photosynth. Res. 41, 357^370.
dation of DD to DT, but partially inhibits the rate of the
[11] Mu«ller, P., Li, X.P. and Niyogi, K.K. (2001) Plant Physiol. 125,
epoxidation reaction. The comparison of £uorescence and pig-
M. Bertrand et al./FEBS Letters 508 (2001) 153^156
[12] Bertrand, M. and Schoefs, B. (1997) in: Handbook of Photosyn-
[20] Rees, D., Noctor, G.D. and Horton, P. (1990) Photosynth. Res.
thesis (Pessarakli, M., Ed.), pp. 151^173, Marcel Dekker, New
[21] Horton, P. and Ruban, A.V. (1992) Photosynth. Res. 34, 375^
[13] Schoefs, B., Bertrand, M. and Funk, C. (2000) Photochem. Pho-
[22] Havaux, M. and Niyogi, N.N. (1999) Proc. Natl. Acad. Sci. USA
[14] Schoefs, B. and Bertrand, M. (2000) FEBS Lett. 486, 243^246;
[23] Lohr, M. and Wilhelm, C. (1999) Proc. Natl. Acad. Sci. USA 96,
[15] Torres, E., Cid, A., Herrero, C. and Abalde, J. (1998) Bioresour.
[24] Yamamoto, H.Y. and Kamite, L. (1972) Biochim. Biophys. Acta
[16] Demmig-Adams, B., Adams, W.W., Heber, U., Meimainis, S.,
Winter, K., Kru«ger, A., Czygan, F.C., Bilger, W. and Bjo«rkman,
[25] Siefermann-Harms, D., Michel, J.-M. and Collard, F. (1980)
O. (1990) Plant Physiol. 92, 292^301.
Biochim. Biophys. Acta 589, 315^323.
[17] Arsalane, W., Rousseau, B. and Duval, J.C. (1994) Photochem.
[26] Pfu«ndel, E.E. and Dilley, R.A. (1993) Plant Physiol. 101, 65^
[18] Jakob, T., Goss, R. and Wilhelm, C. (1999) Plant Biol. 1, 76^82.
[27] Jakob, T., Goss, R. and Wilhelm, C. (2001) J. Plant Physiol. 158,
[19] Genty, B., Wonders, J. and Baker, N.R. (1990) Photosynth. Res.
Condensed by Berni Michel-Samuels, CBC Administrative Assistant SPECIAL SESSION on February 3rd, 2005 10-signature Approve grant application for funding from US Department 2005-038.com of Commerce, Economic Development Administration in the HM, GD, DL, MH) 0 AGAINST 0 amount of $43,000 as submitted by the Tribal Planning Department. The Chair or designee is authorized to sign. 10-s
Haben Sie alles, was Sie für die Pflege Ihres kranken Kindes benötigen? In der nachfolgenden Liste finden Sie das, was Ihre Haus- bzw. Reiseapotheke enthalten sollte. Zeigen Sie Ihre Hausapotheke auch den Personen, die Ihre Kinder hüten. Medikamente / wichtige Utensilien Digitaler Fiebermesser (Achtung: bei Kindern unter 2 Jahren Temperatur immer rektal messen). Bei Fieber und Schmer
 Cadmium inhibits epoxidation of diatoxanthin to diadinoxanthin in the
xanthophyll cycle of the marine diatom Phaeodactylum tricornutum
Martine BertrandaYbY*, Beno|ªt SchoefsbY1, Pavel Si¡elc, Karel Rohacekc, Istvan Molnarc
aInstitut National des Sciences et Techniques de la Mer, Conservatoire National des Arts et Me¨tiers, BP 324, F-50103 Cherbourg Cedex, France
bLaboratory of Biomembranes, University of South Bohemia, Branisovska 31, CZ-370 05 Ceske Budejovice, Czech Republic
cLaboratory of Photosynthesis, Institute of Plant Molecular Biology, Academy of Sciences of the Czech Republic, Branisovska 31,
CZ-370 05 Ceske Budejovice, Czech Republic
Received 6 August 2001; accepted 4 October 2001
Abstract Cd has pleiotropic effects on plant physiology and in
higher plants and green algae, the xanthophyll cycle consists
particular on photosynthesis. It has not been established yet if Cd
in the reversible conversion of violaxanthin to zeaxanthin via
alters the functioning of the xanthophyll cycle. To answer this
antheraxanthin, whereas in chromophytes, it consists in the
question, an exponentially growing culture of the marine diatom
interconversion of diadinoxanthin (DD) and diatoxanthin
Phaeodactylum tricornutum was incubated with Cd (20 mg/l) for
(DT) ([9], reviewed in [8]). The activity of this cycle has
24 h and irradiated with a light activating the xanthophyll cycle,
been correlated with the thermal dissipation of the excess en-
which in diatoms, consists of the reversible deepoxidation of
ergy at the antenna side of the photosynthetic apparatus
diadinoxanthin to diatoxanthin. The measurements show that the
deepoxidation step is not influenced by Cd. In contrast, the Cd
In this contribution we report evidence, based on absor-
concentration used sharply inhibits the epoxidation of diatox-
bance and £uorescence measurements and pigment quanti¢-
anthin to diadinoxanthin. ß 2001 Federation of European Bio-
chemical Societies. Published by Elsevier Science B.V. All
cations, for the partial inhibition by Cd of the epoxidation
step of the xanthophyll cycle in the marine diatom Phaeodac-
Key words: Algae; Microalgae; Cd; Heavy metal stress;
The experiments were performed with the marine diatom P. tricor-
nutum BOHLIN COUGHLAN/-632 axenic strain from the collection
at the Institute of Botany at Trebon, Czech Republic. The cells were
There is considerable concern about the increasing levels of
grown in a medium (pH 8.3) containing ¢ltered reconstituted sea
heavy metals in the environment and the potential harmful
water (33 g/l of commercial salts from the Red Sea), Na2HPO4 (140
WM), NaNO3 (1560 WM), Na2SiO3 (130 WM), CuSO4 (1.4 WM), Co-
e¡ects of these elements on living organisms. This is especially
(NO3)2W6H2O (1.6 WM), MnCl2 (3.2 WM), Mo (0.7 WM), ZnCl2 (0.3
important in the case of heavy metals like Cd, which have no
WM), Fe^EDTA (100 WM), vitamin B1 (54 WM), vitamin B12 (0.7 WM)
and vitamin H (700 WM). The chemicals (P.A. quality) were purchased
The photosynthetic process is very sensitive to Cd: it re-
by Fluka, and the water was of Milli-Q quality (Millipore).
Cadmium inhibits epoxidation of diatoxanthin to diadinoxanthin in the
xanthophyll cycle of the marine diatom Phaeodactylum tricornutum
Martine BertrandaYbY*, Beno|ªt SchoefsbY1, Pavel Si¡elc, Karel Rohacekc, Istvan Molnarc
aInstitut National des Sciences et Techniques de la Mer, Conservatoire National des Arts et Me¨tiers, BP 324, F-50103 Cherbourg Cedex, France
bLaboratory of Biomembranes, University of South Bohemia, Branisovska 31, CZ-370 05 Ceske Budejovice, Czech Republic
cLaboratory of Photosynthesis, Institute of Plant Molecular Biology, Academy of Sciences of the Czech Republic, Branisovska 31,
CZ-370 05 Ceske Budejovice, Czech Republic
Received 6 August 2001; accepted 4 October 2001
Abstract Cd has pleiotropic effects on plant physiology and in
higher plants and green algae, the xanthophyll cycle consists
particular on photosynthesis. It has not been established yet if Cd
in the reversible conversion of violaxanthin to zeaxanthin via
alters the functioning of the xanthophyll cycle. To answer this
antheraxanthin, whereas in chromophytes, it consists in the
question, an exponentially growing culture of the marine diatom
interconversion of diadinoxanthin (DD) and diatoxanthin
Phaeodactylum tricornutum was incubated with Cd (20 mg/l) for
(DT) ([9], reviewed in [8]). The activity of this cycle has
24 h and irradiated with a light activating the xanthophyll cycle,
been correlated with the thermal dissipation of the excess en-
which in diatoms, consists of the reversible deepoxidation of
ergy at the antenna side of the photosynthetic apparatus
diadinoxanthin to diatoxanthin. The measurements show that the
deepoxidation step is not influenced by Cd. In contrast, the Cd
In this contribution we report evidence, based on absor-
concentration used sharply inhibits the epoxidation of diatox-
bance and £uorescence measurements and pigment quanti¢-
anthin to diadinoxanthin. ß 2001 Federation of European Bio-
chemical Societies. Published by Elsevier Science B.V. All
cations, for the partial inhibition by Cd of the epoxidation
step of the xanthophyll cycle in the marine diatom Phaeodac-
Key words: Algae; Microalgae; Cd; Heavy metal stress;
The experiments were performed with the marine diatom P. tricor-
nutum BOHLIN COUGHLAN/-632 axenic strain from the collection
at the Institute of Botany at Trebon, Czech Republic. The cells were
There is considerable concern about the increasing levels of
grown in a medium (pH 8.3) containing ¢ltered reconstituted sea
heavy metals in the environment and the potential harmful
water (33 g/l of commercial salts from the Red Sea), Na2HPO4 (140
WM), NaNO3 (1560 WM), Na2SiO3 (130 WM), CuSO4 (1.4 WM), Co-
e¡ects of these elements on living organisms. This is especially
(NO3)2W6H2O (1.6 WM), MnCl2 (3.2 WM), Mo (0.7 WM), ZnCl2 (0.3
important in the case of heavy metals like Cd, which have no
WM), Fe^EDTA (100 WM), vitamin B1 (54 WM), vitamin B12 (0.7 WM)
and vitamin H (700 WM). The chemicals (P.A. quality) were purchased
The photosynthetic process is very sensitive to Cd: it re-
by Fluka, and the water was of Milli-Q quality (Millipore).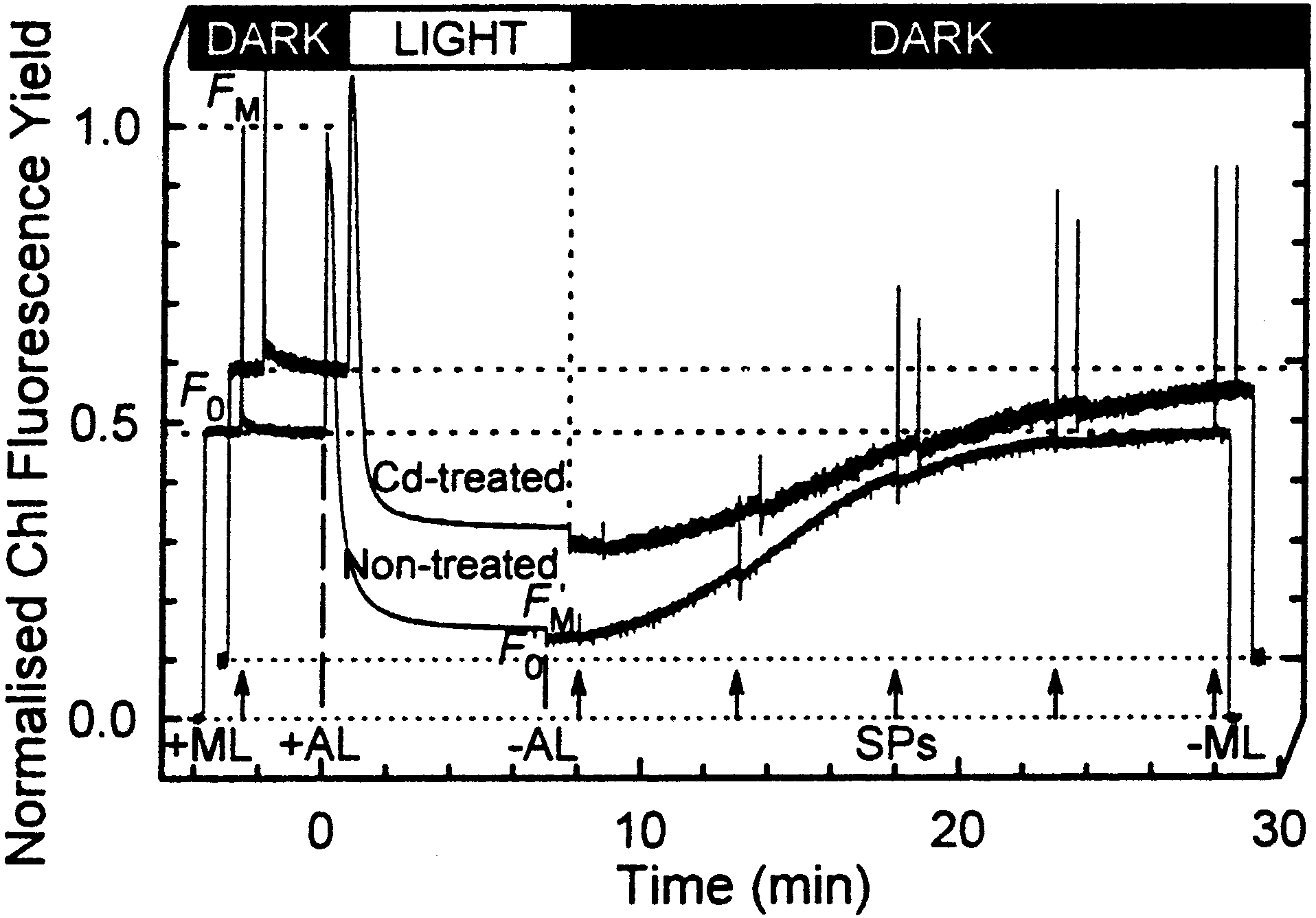
 M. Bertrand et al./FEBS Letters 508 (2001) 153^156
nated with AL (1000 Wmol/m2/s, light source: KL 1500 lamp, Walz,
Germany) for 7 min to induce £uorescence quenching. The intensity
of AL was high enough to keep the PSII closed throughout the ex-
posure and thus the observed £uorescence decline only represents
When AL was switched o¡ and FR was applied, the minimum Chl
£uorescence level corresponding to the light adapted state (F0P) was
determined. Then, dark relaxation of both F0P and maximum Chl
£uorescence level of the light-adapted state (FMP) were measured.
M. Bertrand et al./FEBS Letters 508 (2001) 153^156
nated with AL (1000 Wmol/m2/s, light source: KL 1500 lamp, Walz,
Germany) for 7 min to induce £uorescence quenching. The intensity
of AL was high enough to keep the PSII closed throughout the ex-
posure and thus the observed £uorescence decline only represents
When AL was switched o¡ and FR was applied, the minimum Chl
£uorescence level corresponding to the light adapted state (F0P) was
determined. Then, dark relaxation of both F0P and maximum Chl
£uorescence level of the light-adapted state (FMP) were measured.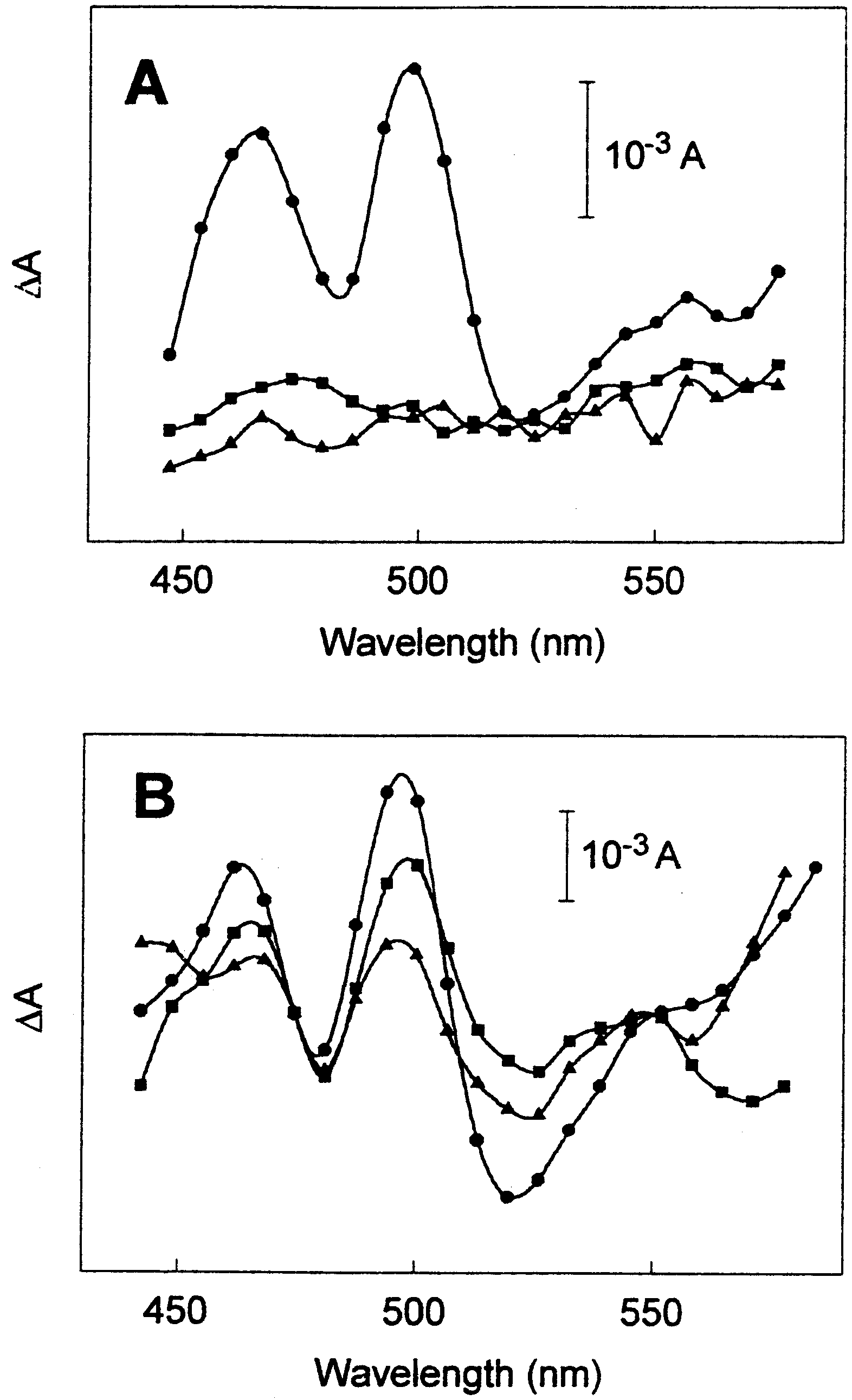 M. Bertrand et al./FEBS Letters 508 (2001) 153^156
er plants, and related £uorescence decrease, usually relax
within 1 min after switching o¡ AL. For Phaeodactylum cells,
the relaxation lasted 10^30 min (Fig. 1). This duration corre-
sponds to the rate of DT epoxidation [23]. Hence, the decrease
in the rate of F0 relaxation, observed in the presence of Cd,
may re£ect an inhibition of the xanthophyll epoxidation re-
3.3. Inhibition of the epoxidation step of the xanthophyll cycle
In order to determine whether the slowing down of the F0
relaxation rate in Cd-treated cells is correlated with a change
in the xanthophyll cycle activity, the previous experiment was
repeated and the variations of the DD and DT amounts were
measured after HPLC separation. Fig. 2 presents the varia-
tions of the relative amount of DT during the light^dark se-
quence in the absence or in the presence of Cd. Regardless of
the presence of Cd, the same level of deepoxidation was
reached at the end of the high-light illumination. In contrast,
the reversion of the xanthophyll cycle was strongly slowed
down in the presence of Cd. Remarkably, the inhibition of
epoxidation rate was very similar in magnitude (approxi-
mately 60%) to that of the relaxation rate (Table 1). This
de¢nitively con¢rms that both phenomenons are tightly
3.4. Measurements of the xanthophyll cycle in vivo
It has been shown with higher plants that the deepoxidation
of violaxanthin to zeaxanthin is re£ected by an increase of the
absorbance at approximately 505 nm [24^26]. In order to
Fig. 3. E¡ect of Cd on the absorbance di¡erence spectra of P. tri-
cornutum (the control stayed in darkness). A: (b) 3Cd+7 min AL;
verify whether the activity of the xanthophyll cycle in diatoms
(F) 3Cd+7 min AL+40 min darkness; (R) 3Cd+DTT+7 min AL.
M. Bertrand et al./FEBS Letters 508 (2001) 153^156
er plants, and related £uorescence decrease, usually relax
within 1 min after switching o¡ AL. For Phaeodactylum cells,
the relaxation lasted 10^30 min (Fig. 1). This duration corre-
sponds to the rate of DT epoxidation [23]. Hence, the decrease
in the rate of F0 relaxation, observed in the presence of Cd,
may re£ect an inhibition of the xanthophyll epoxidation re-
3.3. Inhibition of the epoxidation step of the xanthophyll cycle
In order to determine whether the slowing down of the F0
relaxation rate in Cd-treated cells is correlated with a change
in the xanthophyll cycle activity, the previous experiment was
repeated and the variations of the DD and DT amounts were
measured after HPLC separation. Fig. 2 presents the varia-
tions of the relative amount of DT during the light^dark se-
quence in the absence or in the presence of Cd. Regardless of
the presence of Cd, the same level of deepoxidation was
reached at the end of the high-light illumination. In contrast,
the reversion of the xanthophyll cycle was strongly slowed
down in the presence of Cd. Remarkably, the inhibition of
epoxidation rate was very similar in magnitude (approxi-
mately 60%) to that of the relaxation rate (Table 1). This
de¢nitively con¢rms that both phenomenons are tightly
3.4. Measurements of the xanthophyll cycle in vivo
It has been shown with higher plants that the deepoxidation
of violaxanthin to zeaxanthin is re£ected by an increase of the
absorbance at approximately 505 nm [24^26]. In order to
Fig. 3. E¡ect of Cd on the absorbance di¡erence spectra of P. tri-
cornutum (the control stayed in darkness). A: (b) 3Cd+7 min AL;
verify whether the activity of the xanthophyll cycle in diatoms
(F) 3Cd+7 min AL+40 min darkness; (R) 3Cd+DTT+7 min AL.