Appl Microbiol Biotechnol (2000) 54: 792±798
M. Saayman á H. J. J. van Vuuren á W. H. van Zyl
M. Viljoen-BloomDifferential uptake of fumarate by Candida utilis
Received: 15 March 2000 / Received revision: 4 July2000 / Accepted: 9 July2000
Abstract The dicarboxylic acid fumarate is an impor- of the TCA cycle (Barnett and Kornberg 1960). Previous
tant intermediate in cellular processes and also serves as studies have shown that L-malate can be utilised by
a precursor for the commercial production of ®ne Candida utilis (CaÂssio and LeaÄo 1993), Candida sphaer-
chemicals such as L-malate. Yeast species dier re- ica (CoÃrte-Real et al. 1989), Hansenula anomala (CoÃrte-
markablyin their abilityto degrade extracellular di- Real and LeaÄo 1990) and Kluyveromyces marxianus
carboxylic acids and to utilise them as their only source (Queiros et al. 1998) as their onlysource of carbon and
of carbon. In this studywe have shown that the yeast energy. In these species, the dissociated form of L-malate
Candida utilis eectivelydegraded extracellular fumarate is transported across the plasma membrane bya
and L-malate, but glucose or other assimilable carbon H+-symport system that is substrate-inducible and
sources repressed the transport and degradation of these subject to glucose repression. In contrast, Schizosac-
dicarboxylic acids. The transport of both dicarboxylic charomyces pombe and Zygosaccharomyces bailii can
acids was shown to be stronglyinducible byeither degrade L-malate onlyin the presence of an assimilable
fumarate or L-malate while kinetic studies suggest that carbon source (Rodriguez and Thornton 1990; Osothsilp
the two dicarboxylic acids are transported by the same and Subden 1986a). Other yeasts such as Saccharomyces
transporter protein. In contrast, Schizosaccharomyces cerevisiae can import L-malate and other dicarboxylic
pombe eectivelydegraded extracellular L-malate, but acids onlyvia simple diusion (Salmon 1987) and is
not fumarate, in the presence of glucose or other as- therefore unable to eectivelydegrade or utilise extra-
similable carbon sources. The Sch. pombe malate cellular L-malate.
transporter was unable to transport fumarate, although
In Sch. pombe, the dissociated form of L-malate is
fumarate inhibited the uptake of L-malate.
activelytransported via a H+-symport system that op-
erates constitutively, whereas the undissociated acid
enters the cell via simple diusion (Baranowski and
Radler 1984; Osothsilp and Subden 1986b; Sousa et al.
1992). The dicarboxylic acids fumarate, D-malate,
succinate, oxaloacetate, maleate, malonate and a-keto-
The C4-dicarboxylic acid fumarate serves as an inter- glutarate acted as competitive inhibitors for the uptake
mediate of the tricarboxylic acid (TCA) cycle that allows of L-malate (Sousa et al. 1992), suggesting a common
for the metabolic ¯ow of carbon between various met- transporter for the uptake of dicarboxylic acids in ®ssion
abolic pathways. Yeast species dier remarkably in their yeast. However, Grobler et al. (1995) showed that
abilityto transport and utilise one or more intermediates L-malate, succinate and malonate, but not a-ketogluta-
rate, were activelytransported bySch. pombe cells.
In addition to its role in metabolic processes, fuma-
rate is also an important precursor for the commercial
M. Saayman á W. H. van Zyl á M. Viljoen-Bloom (&)
Department of Microbiology, University of Stellenbosch,
production of ®ne chemicals such as L-malate. The D,L-
Private Bag X1, Matieland 7602, South Africa
malate racemic mixture is routinelyused in a varietyof
foods and beverages whereas the L-isomer is used for the
treatment of conditions such as hyperammonaemia
(Rosenberg et al. 1999). The racemic mixture is com-
merciallyproduced via chemical hydratation of maleate
B.C. Wine Research Centre, Facultyof Agricultural Sciences,
Universityof British Columbia, Vancouver BC,
or fumarate, and the L-isomer through the enzymatic
conversion of fumarate using fumarase-containing
microbial cells. The bioconversion of fumarate to L-
malate has been obtained bystrains of Brevibacterium
(Takata et al. 1980), Candida rugosa (Yang et al. 1992),
Pichia (Keruchen'ko et al. 1995) and Dipodascus (Ro-
senberg et al. 1999) that exhibit high fumarase activities.
Over-expression of the Sac. cerevisiae fumarase gene,
FUM1, also resulted in an increased conversion rate of
fumarate to L-malate (Peleg et al. 1990). Since Sac. ce-
revisiae can onlyimport fumarate through diusion, the
introduction of a fumarate transporter gene into Sac.
cerevisiae could enable this yeast to actively transport
fumarate and consequentlyimprove the bioconversion
Heterologous expression of the Sch. pombe malate
transporter gene, mae1, in a strain of Sac. cerevisiae
resulted in the active transport and ecient degradation
L-malate (Volschenk et al. 1997a, b). Our ®rst ap-
proach was therefore to determine whether expression of
the mae1 gene in Sac. cerevisiae would also enable the
recombinant strain to transport fumarate. We found
that neither the recombinant Sac. cerevisiae strain nor
the wild type Sch. pombe strain could transport fuma-
rate. In search of an alternative fumarate transporter,
several yeast species were evaluated for their ability to
degrade extracellular fumarate. Since C. utilis proved to
be able to degrade both fumarate and L-malate, the
transport of these dicarboxylic acids was further inves-
The yeast strains used in the transport studies included C. utilis
ATCC 9950 T, Sch. pombe 972 h) (Osothsilp 1987), Sch. pombe
972 h) leu1-32 TR) mae1) (Osothsilp and Subden 1986b) and Sac.
cerevisiae YPH259 (MATa ura3-52, lys2-801a, ade2-101o, his3D200,
leu2-D1) (Sikorski and Hieter 1989). The strains used for the screen
on fumarate/malate indicator plates are listed in Table 1. Unless
otherwise stated, the growth media contained 0.17% YNB (yeast
nitrogen base without amino acids and ammonium sulphate [Difco
with amino acids and buered at pH 3.5. Dierent concentrations
of fumarate, L-malate and/or a carbon source were added as indi-
cated for the dierent experiments.
Degradation of extracellular fumarate and
Indicator agar plates were used to screen dierent yeast species for
the degradation of fumarate and L-malate in the presence of dif-
ferent carbon sources. The yeast strains were streaked onto YNB
agar plates containing 0.05% bromocresol-green, 0.3% fumarate
or L-malate, together with 2% glucose, fructose, galactose, glyc-
erol, maltose, ranose or sucrose as carbon source. The plates were
incubated at 30 °C for 2 days and evaluated for a colour change
from yellow at pH 3.3 to blue at pH 5.2.
The utilisation of extracellular fumarate and
C. utilis and Sch. pombe was determined after growth in liquid
YNB media containing 2% glucose, ranose or glycerol/ethanol
as carbon source, supplemented with either 0.5% fumarate or
malate. Cells were harvested at dierent time intervals and high
performance liquid chromatography(HPLC) was used to deter-
mine the residual levels of fumarate and L-malate. Glucose con-
centrations were measured with the glucose oxidase method
(Glucose [Trinder], Sigma, St Louis, Mo.) and cell growth was
determined spectrophotometricallyat OD600. All assays were done Results
Cells of Sac. cerevisiae YPH259 transformed with plasmid pHV3
Strains of Sac. cerevisiae cannot transport extracellular
containing the Sch. pombe mae1 gene (Volschenk et al. 1997b), and
wild type Sch. pombe 972 cells were grown in YNB media con-
dicarboxylic acids such as L-malate or fumarate (Salmon
taining 2% glucose. For C. utilis, cells were cultured in 0.5%
1987). However, transport studies with a recombinant
fumarate, 0.5% L-malate, 2% glucose, 2% ranose or 2% glyc-
Sac. cerevisiae strain expressing the mae1 gene of Sch.
erol/ethanol as the onlysource of carbon. To further investigate the
pombe, showed that active transport of 14C-malate was
eect of dierent carbon sources on the transport of fumarate and
obtained, whereas fumarate was not transported
L-malate, C. utilis cells were cultured to OD600 of 0.6 in media
(Fig. 1a). The active transport of 14C-malate bythe wild
glycerol/ethanol and divided into two batches. One batch of cul-
type Sch. pombe 972 h) strain was con®rmed (Fig. 1b),
tures was assayed immediately while the other was transferred to
but not in the Sch. pombe mae1) mutant strain that has
fresh medium containing either 0.5% L-malate or 2% glucose as
a defective malate transporter. No transport of 14C-
carbon source and incubated for another 6 h.
Cells were harvested in the exponential growth phase (OD
fumarate was observed in either strain (Fig. 1b). How-
0.6), washed twice with ice-cold distilled water and resuspended in
ever, increasing concentrations of fumarate progres-
0.1 M KH2PO4 (pH 3.5) to a ®nal concentration of approximately sivelyinhibited L-malate uptake bythe recombinant Sch.
20 mg dryweight ml)1 (adapted from Grobler et al. 1995). Cell
pombe malate transporter (Fig. 1c). The HPLC analyses
suspensions were pre-incubated for 5 min at 30 °C in a shaker
waterbath at 100 rpm. Assays were initiated by adding 10 ll of
con®rmed that Sch. pombe cells removed a signi®cant
an aqueous solution of [1-14C]-fumarate (6.62 lCi/lmol; ICN
portion (approximately65%) of the L-malate from the
Pharmaceuticals, CA) or L-[1,4(2,3)-14C]-malate (55 lCi/lmol;
glucose-containing growth media within 28 h (Fig. 2a),
Amersham, Bucks, UK). Non-speci®c binding of 14C-fumarate or
whereas less than 15% of the fumarate was removed
14C-malate to the yeast cells was determined by pre-boiling the cells (Fig. 2b). Similar results were obtained for cells grown
for 5 min at 100 °C. Samples of 0.5 ml were withdrawn at dierent
time intervals and the reactions were stopped bydilution with 5 ml
in media containing ranose or glycerol/ethanol as
ice-cold distilled water. The cells were rapidly®ltered through
0.45 lm membranes (Millipore Corporation, Bedford, Mass.) and
Since fumarate inhibited the transport of L-malate in
immediatelywashed with 5 ml ice-cold distilled water. The ®lters
the recombinant Sac. cerevisiae strain without being
were air dried for 10 min and placed in scintillation vials with 5 ml
scintillation reaction mixture (EcoLite, ICN Pharmaceuticals,
transported itself, the uptake and subsequent cellular
Calif.). Levels of radioactivitywere measured with a Beckman
localisation of 14C-malate and fumarate was further in-
LS 3801 scintillation counter (Beckman Instruments, Calif.).
vestigated in wild type Sch. pombe cells (Table 2). An
hour after the addition of 14C-malate to glucose-grown
Cellular fractionation for localisation of dicarboxylic acids
cells, approximately30% of the 14C-malate was re-
moved from the extracellular fraction. The majorityof
Cultures of Sch. pombe in 10 ml YNB medium containing 2%
this was alreadyfurther metabolised to pyruvate and
glucose were harvested at OD600 of 0.6 and resuspended in 1 ml of CO
2 with only0.36% and 0.32% retained in the cell
2PO4 (pH 3.5). Cultures were incubated for another hour
with 1 ll of either 14C-fumarate (6.62 lCi/lmol) or 14C-malate
debris and intracellular fractions, respectively. The ad-
(55 lCi/lmol). A ®nal concentration of 0.5% non-labelled fuma-
dition of unlabelled fumarate decreased the uptake of
L-malate was added to the 14C-malate or 14C-fumarate
C-malate by20% and reduced the localisation of 14C-
cultures, respectively. Cells were harvested and the supernatant was
malate in the cell debris and intracellular fractions by
transferred to scintillation vials containing 5 ml scintillation reac-
tion mixture. The cells were resuspended in 300 ll of 0.1 M
When 14C-fumarate was added to the Sch. pombe
2PO4 (pH 3.5) together with 0.3 g glass beads (106 lm diame-
ter). Cells were broken with 10 pulses of 15 s with 1 min on ice
cells, only1.9% was removed from the extracellular
between pulses. The supernatant and cell debris were separated
fraction after 1 h, but almost 10% of this was retained in
through centrifugation and transferred to scintillation vials con-
the cell debris (Table 2). Although the addition of un-
taining 5 ml scintillation reaction mixture. The levels of radioac-
tivitywere determined as described above.
labelled L-malate did not signi®cantlyin¯uence the up-
take of 14C-fumarate, it decreased its localisation in the
cell debris bymore than 50%. These results suggested
Kinetic parameters for protein-mediated transport
that fumarate competes for the uptake of L-malate by
Cells of C. utilis were cultured in YNB medium containing 0.5%
inhibiting its binding to the malate transporter, although
fumarate as the onlycarbon source. Cells were harvested in the
only L-malate is activelytransported bythe protein.
exponential growth phase (OD600 of 0.6), washed twice with ice-
cold distilled water and resuspended in 0.1 M KH2PO4 (pH 3.5) to
a ®nal concentration of 7 mg dryweight ml)1. Transport assays
were initiated byadding increasing concentrations of 14C-fumarate
Screening of yeast species for degradation
(0.015±2 mM) in the presence or absence of 2 mM non-labelled
L-malate. Estimates of kinetic parameters were obtained from
Lineweaver-Burk plots of the initial uptake rates of 14C-fumarate.
In a screen for yeasts capable of transporting fumarate
m for total dicarboxylic acids was based on the concentra-
tions of both anionic and undissociated dicarboxylic acids.
and L-malate, several yeast species were screened for
Fig. 2 HPLC analyses of extracellular concentrations of a L-malate
and b fumarate during growth of Sch. pombe 972 h) on medium
their abilityto degrade extracellular fumarate or L-
malate incorporated into fumarate/malate indicator agar
plates (Table 1). The yeasts Sch. pombe and Sac. cere-
visiae are not able to utilise intermediates of the TCA
cycle as their only source of carbon (Barnett and
Kornberg 1960), whereas the other species that were
investigated are known for their abilityto utilise TCA
cycle intermediates. No degradation of either fumarate
or L-malate was found for Sac. cerevisiae, since the yeast
is unable to transport either of the dicarboxylic acids. In
Sch. pombe, L-malate was eectivelydegraded in the
presence of all the carbon sources that sustained growth,
but no degradation of fumarate was observed. For
C. utilis, degradation of both fumarate and L-malate
Fig. 1 Transport studies to determine the uptake of L-malate and
were found in all the carbon sources investigated, except
fumarate bystrains of Schizosaccharomyces pombe and Saccharomy-
for glucose (Table 1). The other yeast species showed
ces cerevisiae. a Uptake of 14C-malate and 14C-fumarate in Sac.
varying abilities to utilise fumarate or L-malate that
cerevisiae cel s transformed with the Sch. pombe mae1 gene. b Uptake
of 14C-malate and 14C-fumarate by Sch. pombe 972 h) (wt) and Sch.
seemed to be dependent on the available carbon source.
pombe mae1) (mae1)) grown in 2% glucose. c Competition by
Since the indicator plates onlyprovided limited infor-
fumarate for the transport of 4 mM 14C-malate at pH 3.5 bythe Sac.
mation, further investigation was required to better
cerevisiae YPH259 host strain (control) or transformed with the Sch.
understand the regulatorymechanisms involved in the
pombe mae1 gene. The yeast strains were grown in 2% glucose
without fumarate, or with 10 mM, 20 mM or 30 mM non-labelled
degradation and transport of fumarate and L-malate by
fumarate added simultaneouslywith the 14C-malate
Cellular distribution (% of total 14C added)
Grown in 2% glucose, add fumarate and 14C-malate
Grown in 2% glucose, add L-malate and 14C-fumarate
both fumarate and L-malate is subject to catabolite
When C. utilis cells were pre-cultured in either fum-
Cells of C. utilis eectivelydegraded extracellular fum- arate or L-malate as the onlycarbon source, most of the
arate when grown in YNB medium containing fumarate 14C-fumarate was taken up within 10 s of addition
as the onlycarbon source (Fig. 3a). However, the de- (Fig. 4a). However, the uptake of 14C-fumarate bycells
gradation of fumarate was less ecient when grown in grown on either glucose or ranose as the onlycarbon
the presence of either ranose or glycerol/ethanol as source was almost non-detectable, with onlya small
carbon source, suggesting that other assimilable carbon amount transported bycells grown on glycerol/ethanol.
sources mayresult in catabolite repression of fumarate Similar results were obtained for the transport of L-
transport. In support of this, the degradation of fuma- malate in C. utilis (data not shown), indicating that ac-
rate bycells grown in glucose/fumarate media only tive transport of both fumarate and L-malate was subject
commenced once the glucose had been depleted to substrate induction byeither dicarboxylic acid.
(Fig. 3b). Similar results were obtained for L-malate
The transport of 14C-malate by C. utilis was further
(data not shown), indicating that the degradation of investigated byshifting cultures grown on dierent
carbon sources to fresh medium containing either 0.5%L-malate or 2% glucose (Fig. 4b, c). 14C-malate was
quicklytransported bycells grown on either fumarate orL-malate, but transport ceased when cells were trans-
ferred to glucose-containing medium (Fig. 4b). Cells were
unable to transport 14C-malate when grown on glucose
or glycerol/ethanol medium, not even when 0.5%L-malate was included in the glucose medium (Fig. 4c).
However, cells grown on glucose medium regained their
abilityto transport 14C-malate when transferred to
medium containing L-malate as the onlycarbon source
Preliminarykinetic studies were done to determine
whether C. utilis uses the same transporter protein for
the uptake of fumarate and L-malate. Lineweaver-Burk
plots of the initial rates of uptake of 14C-labelled fum-
arate at pH 3.5 were linear over the concentration range
of 0.08±2 mM (Fig. 5). The following kinetic parameters
were calculated: Vmax(fumarate)(pH 3.5) 1.058 nmol s)1
mg (dryweight) cells)1; Km(pH 3.5) = 0.11 mM. These
results indicated that fumarate and L-malate were mu-
tuallycompetitive inhibitors, suggesting that theymight
share the same carrier protein in C. utilis.
The dicarboxylic acid L-malate is widelyemployed in
both the pharmaceutical and food industries. Due to its
industrial importance, several groups have investigated
the bioconversion of fumarate to L-malate using mi-
crobial cells (Takata et al. 1980; Yang et al. 1992;
Fig. 3 HPLC analyses showing the residual levels of fumarate after
Keruchen'ko et al. 1995; Rosenberg et al. 1999).
growth of C. utilis on a 0.5% fumarate, 2% ranose or 2% glycerol/
ethanol or b 2% glucose as carbon source (residual concentration of
Increased bioconversion of fumarate to L-malate
(80.4 mmol fumaric acid/h per g of cell wet weight) was
Fig. 5 Lineweaver-Burk plots of the initial uptake rates of 4 mM 14C-
fumarate byfumarate-grown cells as a function of the fumarate
concentration in the media. Assays were done in the presence or
obtained byover-expression of the Sac. cerevisiae fu-
marase gene, FUM1 (Peleg et al. 1990). This eciency
maybe further improved if the Sac. cerevisiae cells were
able to activelytransport fumarate and not have to rely
onlyon diusion of the substrate. This could be realised
through heterologous expression of a suitable fumarate
transporter from another yeast in Sac. cerevisiae.
A screen for yeast strains that could degrade extracel-
lular fumarate showed signi®cant dierences in the regu-
lation and speci®cityfor the uptake of fumarate andL-malate between yeast species. A common dicarboxylic
acid transporter was suggested for Sch. pombe strain
ICV'M (Sousa et al. 1992), but results presented here
showed that neither the wild type Sch. pombe 972 h) strain
nor a recombinant strain of Sac. cerevisiae containing the
Sch. pombe malate transporter gene was able to transport
fumarate (Fig. 1). However, increasing concentrations of
fumarate were able to progressivelyinhibit the uptake ofL-malate bythe recombinant strain. Cellular fractiona-
tion of glucose-grown cells (Table 2) showed that the
addition of unlabelled fumarate decreased both the up-
take and membrane localisation of 14C-malate. The data
suggested that fumarate can also bind to the malate
transporter and therefore inhibit the uptake of L-malate.
The binding of both fumarate and L-malate to the Sch.
pombe malate transporter can be ascribed to the struc-
Fig. 4 Transport studies to determine the uptake of 14C-labelled L-
tural relatedness of the two dicarboxylic acids. Similarly,
malate and fumarate by C. utilis. a Uptake of 14C-fumarate after
Grobler et al. (1995) reported that a-ketoglutarate was
growth on 2% glucose, 2% ranose, 2% glycerol/ethanol, 0.5%
fumarate or 0.5% L-malate as onlycarbon source. b Uptake of 14C-
not transported by Sch. pombe, although it competed for
malate after growth on 0.5% fumarate or 0.5% L-malate as only
the uptake of L-malate (Sousa et al. 1992).
carbon source, and shifted to fresh medium containing 2% glucose. c
The results presented here indicate a signi®cant dif-
Uptake of 14C-malate after growth on 2% glucose or 2% glycerol/
ference in the transport of fumarate and L-malate by
ethanol with or without 0.5% L-malate. Glucose-grown cells were also
C. utilis and Sch. pombe. Cells of Sch. pombe 972 eec-
shifted to fresh medium containing 0.5% L-malate
tivelytransported L-malate, but not fumarate, and no
evidence for substrate induction or glucose repression
for the uptake of L-malate was found. In contrast, the
C. utilis ATCC 9950 T strain eectivelytransported
both fumarate and L-malate and the uptake of both References
dicarboxylic acids was induced by either of the sub-
strates. The kinetic data suggest that fumarate and
Baranowski K, Radler F (1984) The glucose-dependent transport
malate are transported bythe same carrier protein in C.
of L-malate in Zygosaccharomyces bailii. Antonie Van Leeu-
utilis, which explains the similar regulatorymechanisms Barnett JA, Kornberg HL (1960) The utilisation byyeast of acids
observed for the transport of fumarate and L-malate.
of the tricarboxylic acid cycle. J Gen Microbiol 23: 65±82
The degradation of either fumarate or L-malate by CaÂssio F, LeaÄo C (1993) A comparative studyon the transport of
C. utilis was sensitive to the presence of glucose (Fig. 3).
l(-)malic acid and other short-chain carboxylic acids in the yeast
Candida utilis: evidence for a general organic acid permease.
This supports previous reports that the utilisation of
L-malate in C. utilis strain IGC 3092 was subject to
CoÃrte-Real M, LeaÄo C (1990) Transport of malic acid and other
glucose repression (CaÂssio and LeaÄo 1993). In addition,
dicarboxylic acids in the yeast Hansenula anomala. Appl Envi-
we observed that the transport of either fumarate or
CoÃrte-Real M, LeaÄo C, Van Uden N (1989) Transport of L-malic
L-malate was also insigni®cant in the presence of other
acid and other dicarboxylic acids in the yeast Candida sphaerica.
carbon sources such as ranose and glycerol/ethanol
(Fig. 4). This con®rmed that C. utilis employs a double Grobler J, Bauer F, Subden RE, Van Vuuren HJJ (1995) The mae1
gene of Schizosaccharomyces pombe encodes a permease for
fumarate with the dicarboxylic acids only being trans-
malate and other C4 dicarboxylic acids. Yeast 11: 1485±1491
Keruchen'ko YS, Kerucheno'ko ID, Gladilin KL (1995) Forma-
ported in the presence of either of the inducers and when
tion of malate byyeast of the genus Pichia. Prikl Biochim
no alternative carbon source is available.
The carbon sensitivityand substrate induction ob- Osothsilp C (1987) Genetic and biochemical studies of malic acid
metabolism in Schizosaccharomyces pombe. PhD dissertation,
C. utilis could be interpreted in the context of its ability Osothsilp C, Subden RE (1986a) Isolation and characterisation of
to utilise intermediates of the TCA cycle as the only
Schizosaccharomyces pombe mutants with defective NAD-
source of carbon and energy. The yeast C. utilis is
dependent malic enzyme. Can J Microbiol 32: 481±486
Crabtree-negative and can therefore ferment sugars only Osothsilp C, Subden RE (1986b) Malate transport in Schizosac-
charomyces pombe. J Bacteriol 168: 1439±1443
under oxygen-limited conditions (Van Dijken et al. Peleg Y, Rokem JS, Goldberg I, Pines O (1990) Inducible overex-
1993). Under aerobic growth conditions, the yeast
pression of the FUM1 gene in Saccharomyces cerevisiae: Lo-
tended to channel most of its pyruvate into the TCA
calization of fumarase and ecient fumaric acid bioconversion
cycle, resulting in an adequate supply of intracellular
to L-malic acid. Appl Environ Microbiol 56: 2777±2783
TCA cycle intermediates such as fumarate and
Queiros O, Casal M, Altho S, Moradas-Ferreira P, LeaÄo C (1998)
Isolation and characterization of Kluyveromyces marxianus
Since the degradation of glucose, ranose or glycerol/
mutants de®cient in malate transport. Yeast 14: 401±407
ethanol can provide pyruvate for the TCA cycle, the Rodriguez SB, Thornton RJ (1990) Factors in¯uencing the utili-
dicarboxylic acids will most likely only be utilised if a
sation of L-malate byyeasts. FEMS Microbiol Lett 72: 17±22
more ecient carbon source is not available. The results Rosenberg M, Mikova H, KrisÏtofõÂkova L (1999) Formation of
presented here support the notion that C. utilis cells al-
low the transport of fumarate and L-malate onlyin the Salmon JM (1987) L-malic acid permeation in resting cells of an-
presence of the inducers and when an alternative carbon
aerobicallygrown Saccharomyces cerevisiae. Biochim Biophys
source is not available. Furthermore, the results pre-
Sikorski RS, Hieter P (1989) A system of shuttle vectors and yeast
sented in Fig. 3 indicate that the catabolite repression is
host strains designed for ecient manipulation of DNA in
stronger when cells are grown on glucose than on the
Saccharomyces cerevisiae. Genetics 122: 19±27
less favourable carbon source glycerol/ethanol.
Sousa MJ, Mota M, LeaÃo C (1992) Transport of malic acid in the
Although the transport of dicarboxylic acids has been
yeast Schizosaccharomyces pombe: evidence for a proton±
described for a number of yeast species, the Sch. pombe
dicarboxylate symport. Yeast 8: 1025±1031
Takata I, Yamamoto K, Tosa T, Chibata I (1980) Immobilization
mae1 gene is the onlymalate transporter gene cloned
of Brevibacterium ¯avum with carrageenan and its application
and sequenced thus far (Grobler et al. 1995). In this
for continuous production of L-malic acid. Microb Technol 2:
study, we demonstrated signi®cant dierences between
Sch. pombe and C. utilis concerning the uptake of fum- Van Dijken JP, Weusthuis RA, Pronk JT (1993) Kinetics of
growth and sugar consumption in yeasts. Antonie Van Leeu-
arate and L-malate and the regulation thereof. However,
a proper investigation into the molecular basis for the Volschenk H, Viljoen M, Grobler J, Bauer F, Lonvaud-Funel A,
transport of fumarate and L-malate by C. utilis can only
Denayrolles M, Subden RE, Van Vuuren HJJ (1997a) Malo-
be done once the fumarate/malate transporter gene from
lactic fermentation in grape musts bya geneticallyengineered
strain of Saccharomyces cerevisiae. Am J Enol Vitic 48: 193±
Volschenk H, Viljoen M, Grobler J, Petzold B, Bauer F, Subden
Acknowledgements We thank Q. Willemse and M. Blom for
RE, Young RA, Lonvaud-Funel A, Denayrolles M, Van
technical assistance with HPLC analyses. This work was funded by
Vuuren HJJ (1997b) Engineering pathways for malate degra-
WINETECH, FRD grant 2016111 and THRIP grant 2038508 to
dation in Saccharomyces cerevisiae. Nat Biotechnol 15: 253±257
H.J.J.v.V., and THRIP grant 2040900 to M.V. Experiments were
Yang LW, Wang XY, Wei S (1992) Immobilization of Candida
conducted in accordance with South African law on the handling of
rugosa having high fumarase activity with polyvinyl alcohol.
nyttjande och avetablering och som inte är trappor, ramper och länkar. Detta utgör av väsentligt annan natur än dessa åtgärder. moduluppställningens begränsningslinje. 1.1 BEGREPPSBESTÄMNINGAR Allmänna avtalsvillkor för modul Etableringsgräns utgörs av färdig iordning uppställning: Bestämmelser enligt detta ställd markyta eller av beställaren tillhandahå
Nutrient-Drug Interactions and Food no. 9.361 by J. Anderson and H. Hart 1 It is a difficult and complex problem to accurately determine theeffects of food and nutrients on a particular drug. There are many dramaticresults or problems caused by food-drug, drug-drug and alcohol-food-druginteractions. The following table is designed to help the reader become moreknowledgeable about drug inter
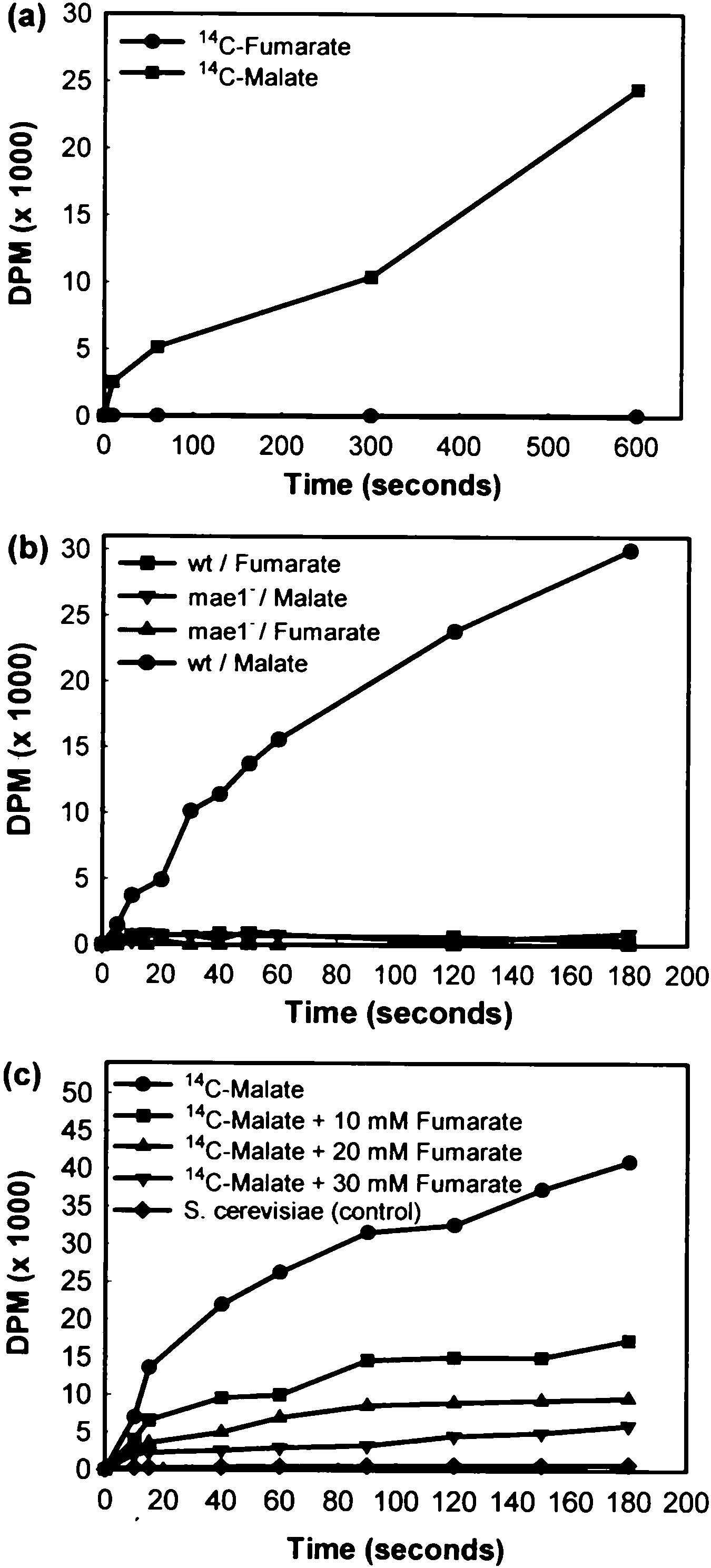
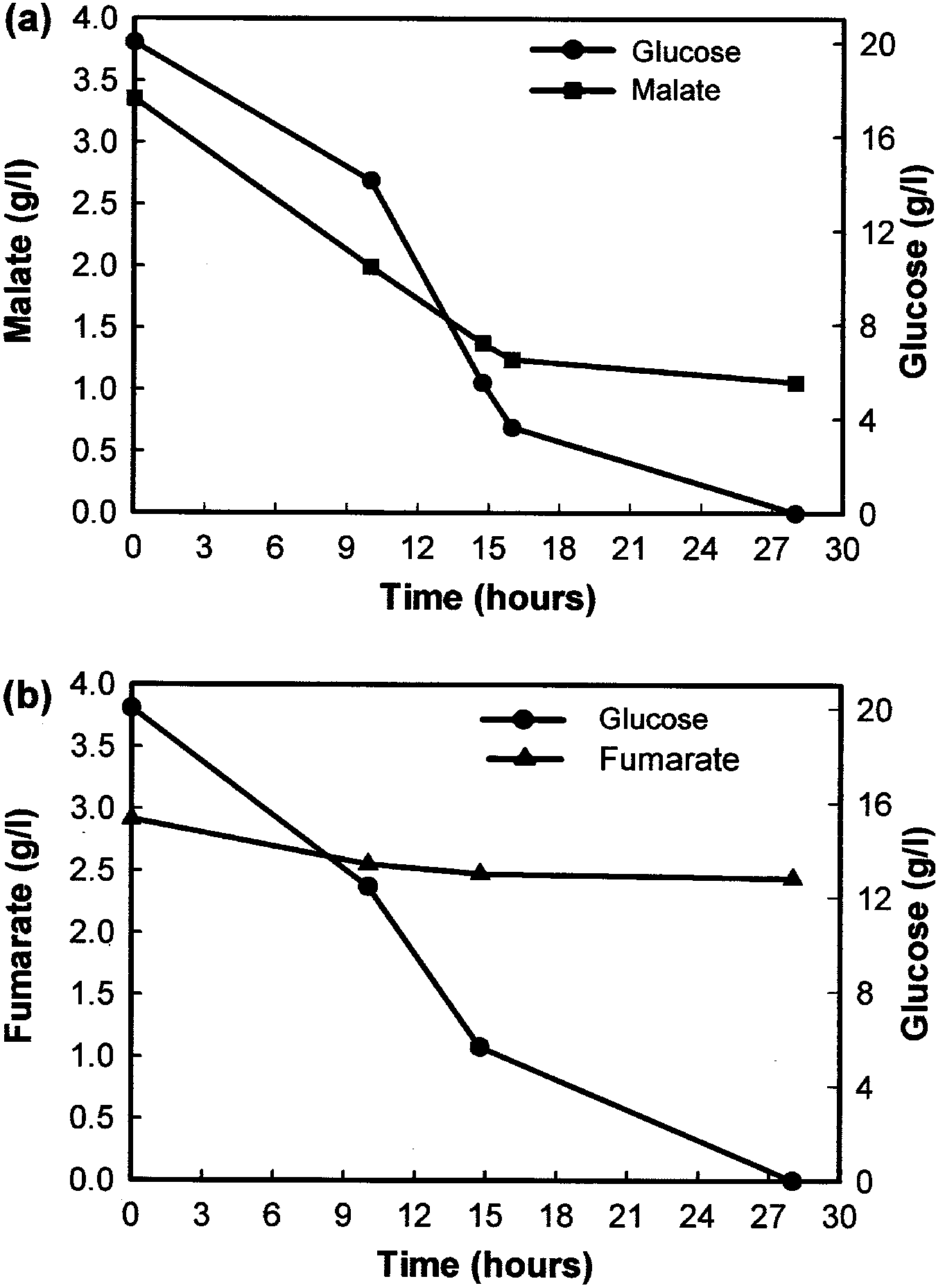 Fig. 2 HPLC analyses of extracellular concentrations of a L-malate
and b fumarate during growth of Sch. pombe 972 h) on medium
their abilityto degrade extracellular fumarate or L-
malate incorporated into fumarate/malate indicator agar
plates (Table 1). The yeasts Sch. pombe and Sac. cere-
visiae are not able to utilise intermediates of the TCA
cycle as their only source of carbon (Barnett and
Kornberg 1960), whereas the other species that were
investigated are known for their abilityto utilise TCA
cycle intermediates. No degradation of either fumarate
or L-malate was found for Sac. cerevisiae, since the yeast
is unable to transport either of the dicarboxylic acids. In
Sch. pombe, L-malate was eectivelydegraded in the
presence of all the carbon sources that sustained growth,
but no degradation of fumarate was observed. For
C. utilis, degradation of both fumarate and L-malate
Fig. 1 Transport studies to determine the uptake of L-malate and
were found in all the carbon sources investigated, except
fumarate bystrains of Schizosaccharomyces pombe and Saccharomy-
for glucose (Table 1). The other yeast species showed
ces cerevisiae. a Uptake of 14C-malate and 14C-fumarate in Sac.
Fig. 2 HPLC analyses of extracellular concentrations of a L-malate
and b fumarate during growth of Sch. pombe 972 h) on medium
their abilityto degrade extracellular fumarate or L-
malate incorporated into fumarate/malate indicator agar
plates (Table 1). The yeasts Sch. pombe and Sac. cere-
visiae are not able to utilise intermediates of the TCA
cycle as their only source of carbon (Barnett and
Kornberg 1960), whereas the other species that were
investigated are known for their abilityto utilise TCA
cycle intermediates. No degradation of either fumarate
or L-malate was found for Sac. cerevisiae, since the yeast
is unable to transport either of the dicarboxylic acids. In
Sch. pombe, L-malate was eectivelydegraded in the
presence of all the carbon sources that sustained growth,
but no degradation of fumarate was observed. For
C. utilis, degradation of both fumarate and L-malate
Fig. 1 Transport studies to determine the uptake of L-malate and
were found in all the carbon sources investigated, except
fumarate bystrains of Schizosaccharomyces pombe and Saccharomy-
for glucose (Table 1). The other yeast species showed
ces cerevisiae. a Uptake of 14C-malate and 14C-fumarate in Sac.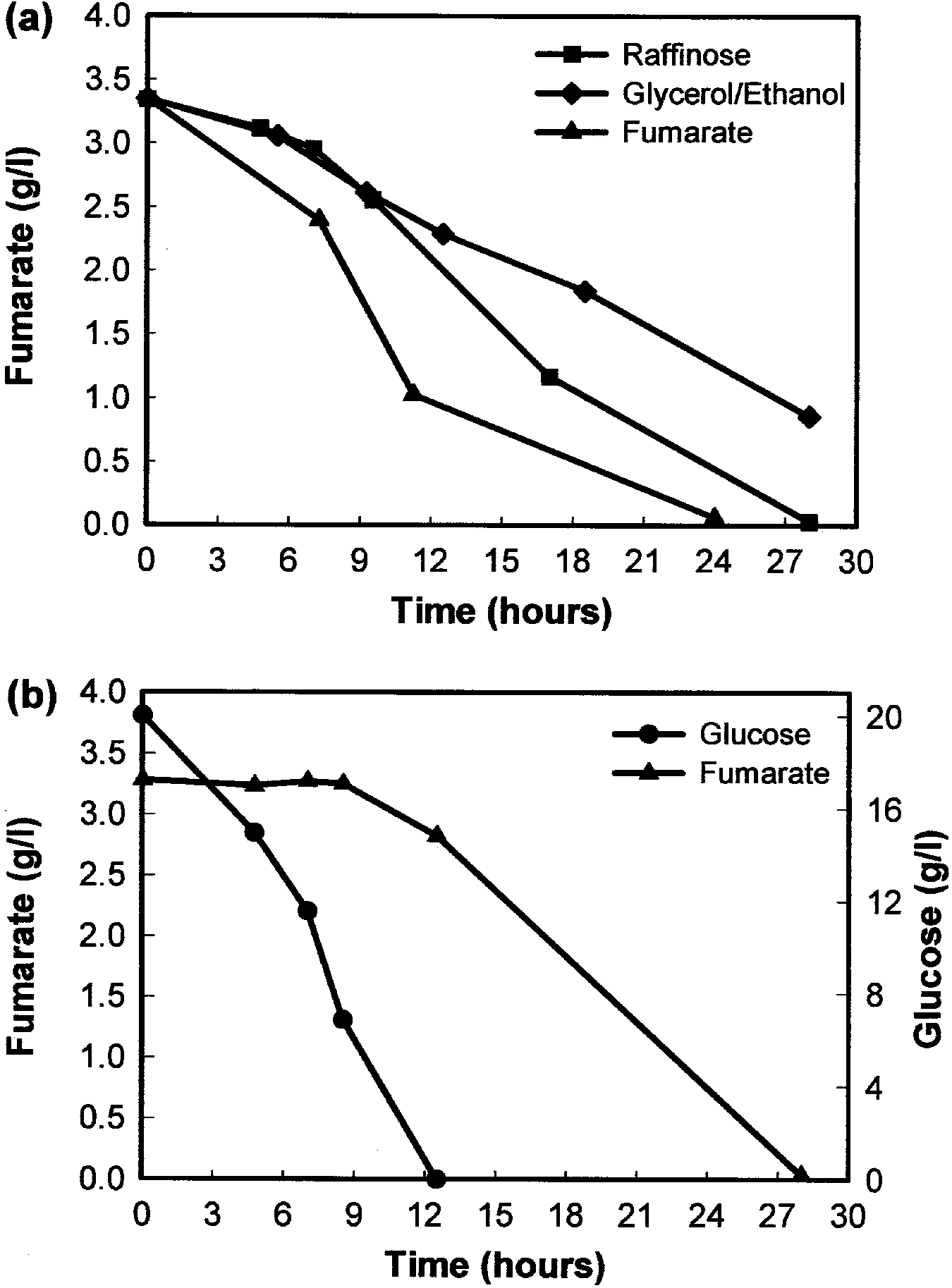 Cellular distribution (% of total 14C added)
Grown in 2% glucose, add fumarate and 14C-malate
Grown in 2% glucose, add L-malate and 14C-fumarate
both fumarate and L-malate is subject to catabolite
When C. utilis cells were pre-cultured in either fum-
Cells of C. utilis eectivelydegraded extracellular fum- arate or L-malate as the onlycarbon source, most of the
arate when grown in YNB medium containing fumarate 14C-fumarate was taken up within 10 s of addition
as the onlycarbon source (Fig. 3a). However, the de- (Fig. 4a). However, the uptake of 14C-fumarate bycells
gradation of fumarate was less ecient when grown in grown on either glucose or ranose as the onlycarbon
the presence of either ranose or glycerol/ethanol as source was almost non-detectable, with onlya small
carbon source, suggesting that other assimilable carbon amount transported bycells grown on glycerol/ethanol.
Cellular distribution (% of total 14C added)
Grown in 2% glucose, add fumarate and 14C-malate
Grown in 2% glucose, add L-malate and 14C-fumarate
both fumarate and L-malate is subject to catabolite
When C. utilis cells were pre-cultured in either fum-
Cells of C. utilis eectivelydegraded extracellular fum- arate or L-malate as the onlycarbon source, most of the
arate when grown in YNB medium containing fumarate 14C-fumarate was taken up within 10 s of addition
as the onlycarbon source (Fig. 3a). However, the de- (Fig. 4a). However, the uptake of 14C-fumarate bycells
gradation of fumarate was less ecient when grown in grown on either glucose or ranose as the onlycarbon
the presence of either ranose or glycerol/ethanol as source was almost non-detectable, with onlya small
carbon source, suggesting that other assimilable carbon amount transported bycells grown on glycerol/ethanol.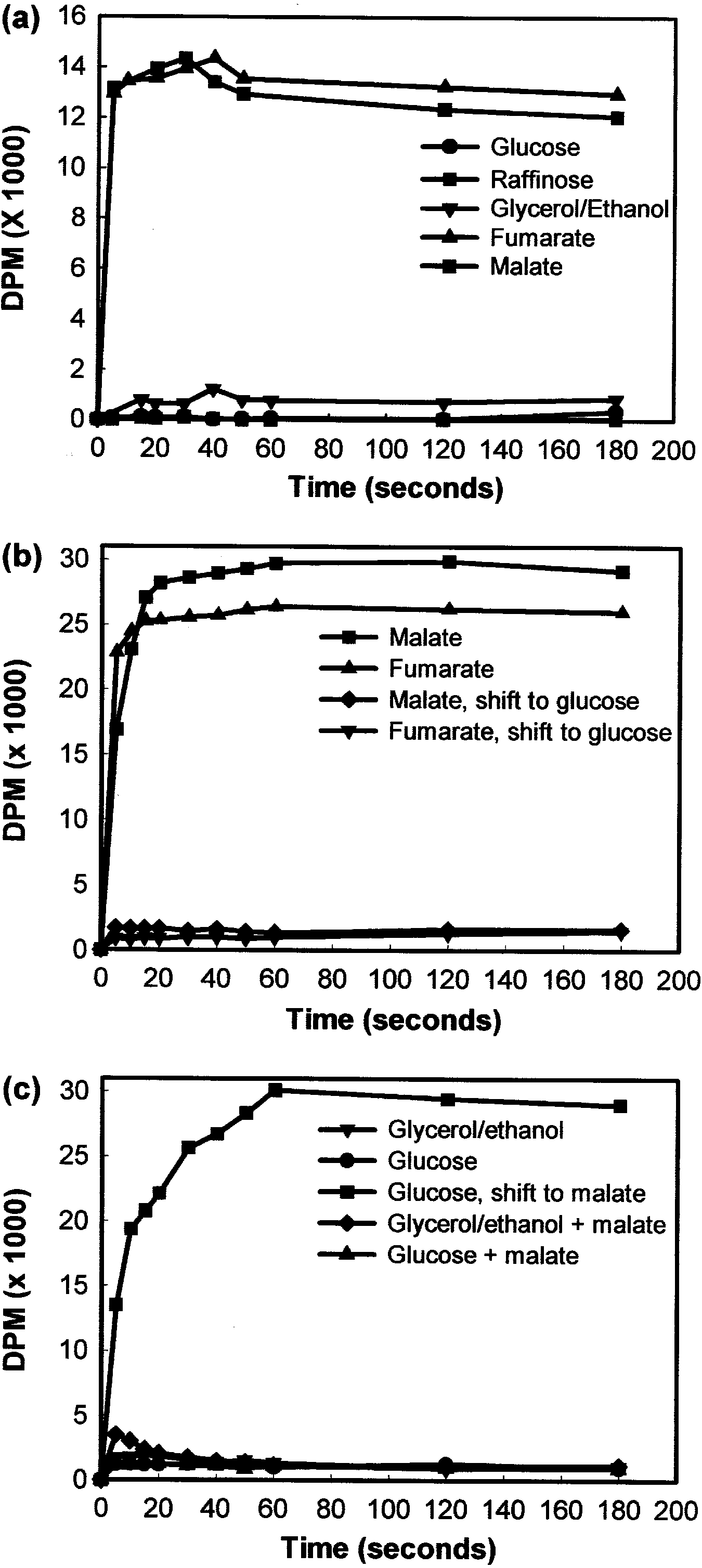
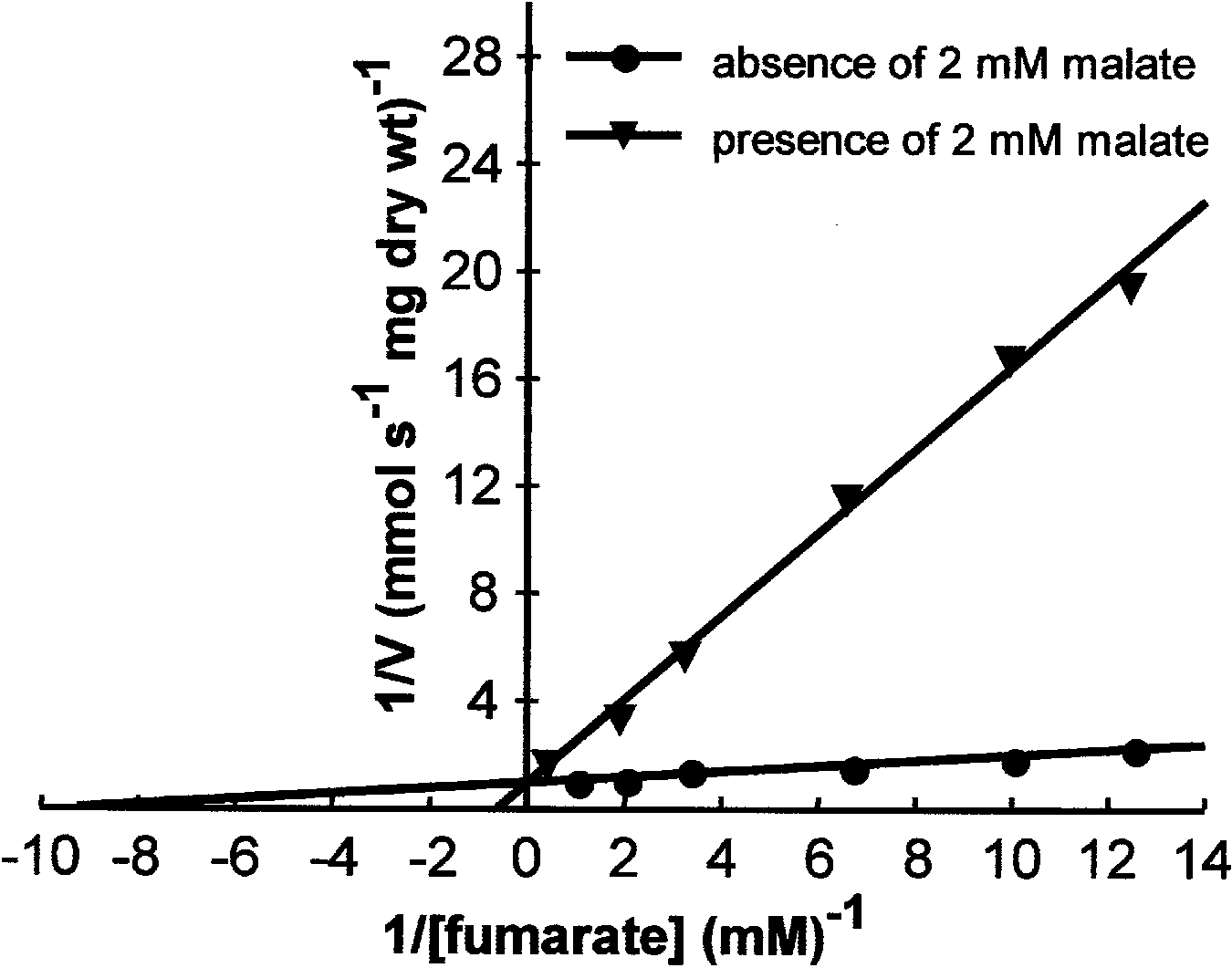 Fig. 5 Lineweaver-Burk plots of the initial uptake rates of 4 mM 14C-
fumarate byfumarate-grown cells as a function of the fumarate
concentration in the media. Assays were done in the presence or
obtained byover-expression of the Sac. cerevisiae fu-
marase gene, FUM1 (Peleg et al. 1990). This eciency
maybe further improved if the Sac. cerevisiae cells were
able to activelytransport fumarate and not have to rely
onlyon diusion of the substrate. This could be realised
through heterologous expression of a suitable fumarate
transporter from another yeast in Sac. cerevisiae.
Fig. 5 Lineweaver-Burk plots of the initial uptake rates of 4 mM 14C-
fumarate byfumarate-grown cells as a function of the fumarate
concentration in the media. Assays were done in the presence or
obtained byover-expression of the Sac. cerevisiae fu-
marase gene, FUM1 (Peleg et al. 1990). This eciency
maybe further improved if the Sac. cerevisiae cells were
able to activelytransport fumarate and not have to rely
onlyon diusion of the substrate. This could be realised
through heterologous expression of a suitable fumarate
transporter from another yeast in Sac. cerevisiae.