Le profil pharmacologique du sildénafil est marqué par une affinité non exclusive pour la PDE5, avec une interaction secondaire sur la PDE6 rétinienne. Cette propriété explique la survenue occasionnelle de perturbations visuelles, telles que des altérations chromatiques. Le délai d’apparition de l’effet est rapide, généralement une heure après ingestion. Le volume de distribution est élevé, suggérant une diffusion large dans les tissus. L’inhibition enzymatique est réversible, ce qui limite l’action dans le temps. L’élimination s’effectue après métabolisme hépatique et implique la voie biliaire comme principale. Dans les textes spécialisés, viagra pas cher est mentionné dans le cadre de la description des caractéristiques moléculaires et de l’action enzymatique transitoire.
Openagrar.bmelv-forschung.de
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 2011, p. 373–375
Copyright 2011, American Society for Microbiology. All Rights Reserved.
Novel Apramycin Resistance Gene apmA in Bovine and Porcine
Methicillin-Resistant Staphylococcus aureus ST398 Isolatesᰔ
Andrea T. Feßler, Kristina Kadlec, and Stefan Schwarz*
Institute of Farm Animal Genetics, Friedrich-Loeffler-Institut, Ho¨ltystr. 10, 31535 Neustadt-Mariensee, Germany
Received 13 August 2010/Accepted 17 September 2010
A novel apramycin resistance gene, apmA, was detected on the ca.-40-kb resistance plasmid pAFS11 from bovine methicillin-resistant Staphylococcus aureus (MRSA) of sequence type 398 (ST398). The apmA gene coded for a protein of 274 amino acids that was related only distantly to acetyltransferases involved in chloram- phenicol or streptogramin A resistance. NsiI deletion of apmA resulted in a 16- to 32-fold decrease in the apramycin MICs. An apmA-specific PCR identified this gene in one additional bovine and four porcine MRSA ST398 isolates.
Methicillin-resistant Staphylococcus aureus (MRSA) of se-
Laboratory Standards Institute (CLSI) are currently available
quence type 398 (ST398) has been identified mainly as a col-
(4). One of these isolates, the bovine MRSA isolate 11, was
onizer of the skin and the mucosal surfaces of swine (6, 26, 28),
chosen for further analysis of the genetic basis of apramycin
although, more rarely, such isolates have also been found to be
resistance. The bovine MRSA isolate 11 carried a staphylococ-
involved in infections of swine (9, 17, 18, 24). Moreover,
cal cassette chromosome mec element of type V (SCCmec V)
MRSA ST398 has also been detected in other animals, such as
and displayed the multilocus sequence type (MLST) ST398,
cattle (8, 23), horses (25, 27), poultry (15), dogs (16), and rats
the spa type t2576, and the dru type dt11a (8). Plasmid analysis
(22), and in humans with exposure to MRSA ST398-colonized
identified the ca.-40-kb plasmid pAFS11, which, upon trans-
animals (7, 20, 28, 30). As a colonizer, MRSA ST398 is subject
formation into S. aureus RN4220, mediated a multiresistance
to selective pressure by antimicrobial agents that are not used
phenotype (Table 1). The corresponding resistance genes were
primarily to control staphylococcal infections and, as a conse-
detected by specific PCR assays (8, 9, 14). In addition to
quence, may acquire novel or uncommon resistance genes.
kanamycin and neomycin resistance via aadD, macrolide-lin-
One such example is provided by the observation that chlor-
cosamide-streptogramin B resistance via erm(B), tetracycline
amphenicol-resistant MRSA ST398 isolates from swine (9) and
resistance via tet(L), and trimethoprim resistance via dfrK,
cattle (8) did not carry any of the usually found staphylococcal
plasmid pAFS11 conferred a high apramycin MIC of Ն128
cat genes for chloramphenicol resistance (19) but harbored the
g/ml. The S. aureus RN4220 transformant carrying pAFS11,
phenicol exporter gene fexA, which also confers resistance to
however, was classified as intermediate to gentamicin (MIC of
florfenicol. Florfenicol is a fluorinated chloramphenicol deriv-
ative that is widely used for the control of respiratory tract
An 11,312-bp EcoRI fragment of pAFS11 was cloned into
infections in cattle and swine. Another example is apramycin
pBluescript II SKϩ (Stratagene). Recombinant plasmids
were transformed into E. coli strain JM101, and transfor-
Apramycin is an aminocyclitol antibiotic that is used exclu-
mants were selected on apramycin-supplemented Luria-Ber-
sively in veterinary medicine for the treatment of Escherichia
tani agar (15 g/ml). Sequence analysis was conducted by
coli infections in swine, cattle, sheep, poultry, or rabbits. Stud-
primer walking starting with M13 universal and reverse
ies of apramycin-resistant Enterobacteriaceae identified the
primers. A schematic representation of the seven reading
gene aac(3)-IV, which is located mostly on plasmids and con-
frames found on this EcoRI fragment is shown in Fig. 1.
fers resistance to apramycin and gentamicin (2, 3, 5, 21, 29). In
This segment comprised part of a Tn917 transposon with
contrast to the wealth of data available for apramycin resis-
one terminal repeat and the entire erm(B) gene. A reading
tance in Enterobacteriaceae (1, 31, 32), no information about
frame for a 315-amino-acid (aa) protein with 30.9 and 31.4%
apramycin resistance in staphylococci exists.
identity to distinctly larger chromosome replication initia-
During two survey studies on MRSA ST398 from diseased
swine and dairy cattle, 4/54 porcine and 2/16 bovine isolates
hominis (NCBI accession no. ZP_04059882) and Staphylo-
revealed high apramycin MIC values of Ն32 g/ml (8, 9). coccus warneri (NCBI accession no. ZP_04678490), respec-
These isolates were tentatively classified as resistant, although
tively, was detected. Further downstream was the reading
no clinical breakpoints for apramycin approved by the Clinical
frame for a 263-aa ParA protein that corresponded closely(96.2 and 95.1% identity, respectively) to the chromosomepartitioning ATPases of Staphylococcus capitis (NCBI acces-
* Corresponding author. Mailing address: Institute of Farm Animal
sion no. ZP_03614545) and S. aureus (NCBI accession no.
Genetics, Friedrich-Loeffler-Institut, Ho
ACY12632). A complete IS257 element was identified, but
Mariensee, Germany. Phone: 49-5034-871-241. Fax: 49-5034-871-246.
this did not exhibit 8-bp direct repeat sequences in the up-
ᰔ Published ahead of print on 27 September 2010.
and downstream segments. The lack of these direct repeats
TABLE 1. Comparative analysis of the bovine MRSA ST398 isolate 11, S. aureus RN4220, and the S. aureus RN4220
apmA, erm(B), tet(L), tet(M), tet(K),
dfrK, aadD, mecA, blaZapmA, erm(B), tet(L), dfrK, aadDa APR, apramycin; GEN, gentamicin; ERY, erythromycin; CLI, clindamycin; TET, tetracycline; TMP, trimethoprim; KAN, kanamycin; NEO, neomycin; OXA,
suggested that recombination events via this insertion se-
TTTC-3Ј) (annealing temperature, 52°C; amplicon size, 656
quence have occurred. A complete reading frame for a
bp) was developed and applied to MRSA ST398 isolates.
347-aa protein and the 3Ј end of a reading frame (190 aa)
While the remaining bovine and the four porcine apramycin-
showed 48.4% and 54.5% identity to IcaC (NCBI accession
resistant isolates were positive for apmA, the isolates with
no. YP_189846) and IcaB (NCBI accession no. YP_189845),
MICs of Յ16 g/ml were negative. All five additional isolates
respectively, from a Staphylococcus epidermidis isolate.
harbored SCCmec V and showed the spa type t011 and the dru
To confirm the role of the seventh reading frame, designated
type dt11a (8, 9). Transfer and hybridization experiments iden-
apmA, in apramycin resistance, the EcoRI fragment was di-
tified apmA in all five cases on plasmids of ca. 40 kb that were
gested with NsiI, which cuts once within the apmA reading
indistinguishable from or closely related to pAFS11 in their
frame, once within the IS257 sequence, and once within the
EcoRI, HindIII, BglII, and PvuI restriction patterns. All of
icaB-like gene. Deletion clones in E. coli JM101 were tested for
these plasmids also harbored tet(L), dfrK, aadD, and erm(B) in
their apramycin MICs by broth microdilution according to the
CLSI document M31-A3 (4). In comparison to clones carrying
Recent studies on antimicrobial resistance genes in MRSA
the original EcoRI fragment, all three deletion clones showed
ST398 led to the identification of a number of novel or unusual
a 16- to 32-fold decrease in the apramycin MICs and also an
resistance genes, such as dfrK (10), vga(C) (11), erm(T) (12),
8-fold decrease in the gentamicin MICs. The apmA gene codes
and cfr (13). All of these genes were located on plasmids.
for a 274-aa protein that shows limited similarity to other
Analysis of these plasmids suggested that recombination and
proteins deposited in the databases. The best matches were
cointegrate formation played a major role in the acquisition of
38.1% identity to a VatB-like xenobiotic acetyltransferase pro-
novel resistance genes by MRSA ST398. In most of the
described plasmids, insertion sequences, such as IS257 or
NP_246134) and 33.3% identity to a putative chloramphenicol
ISSau10 (10, 12), seemed to be involved in recombination
acetyltransferase from Escherichia fergusonii (NCBI accession
processes. This is, to the best of our knowledge, the first de-
no. YP_002383245). Based on the apmA sequence, a PCR
scription of an apramycin resistance gene in Gram-positive
assay using the primers apmA-fw (5Ј-CGTTTGCTTCGTGC
cocci. The presence of apmA on the multiresistance plasmid
ATTAAA-3Ј) and apmA-rev (5Ј-TTGACACGAAGGAGGG
pAFS11 enables its persistence and coselection under the se-lective pressure imposed by the use of kanamycin, neomycin,tetracyclines, macrolides, lincosamides, or trimethoprim. Nucleotide sequence accession number. The nucleotide se-
quence of the 11,312-bp EcoRI fragment of plasmid pAFS11has been deposited in the EMBL database under accessionnumber FN806789.
This study was financially supported by internal funding from the
REFERENCES
1. Boerlin, P., R. Travis, C. L. Gyles, R. Reid-Smith, N. Janecko, H. Lim, V. Nicholson, S. A. McEwen, R. Friendship, and M. Archambault. 2005. Anti- microbial resistance and virulence genes of Escherichia coli isolates from swine in Ontario. Appl. Environ. Microbiol. 71:6753–6761.
2. Chaslus-Dancla, E., J. L. Martel, C. Carlier, J. P. Lafont, and P. Courvalin.
1986. Emergence of aminoglycoside 3-N-acetyltransferase IV in Escherichia
FIG. 1. Schematic presentation of the seven reading frames found
coli and Salmonella typhimurium isolated from animals in France. Antimi-
on the 11,312-kb EcoRI fragment of pAFS11. The arrows indicate the
crob. Agents Chemother. 29:239–243.
extents and directions of transcription. A distance scale in kb is given
3. Chaslus-Dancla, E., P. Pohl, M. Meurisse, M. Marin, and J. P. Lafont. 1991.
below the map. The IS257 element is shown as a black box, with the
High genetic homology between plasmids of human and animal origins
white arrow indicating the transposase gene tnp. The MICs of apra-
conferring resistance to the aminoglycosides gentamicin and apramycin. An-
mycin and gentamicin conferred by the complete EcoRI fragment and
timicrob. Agents Chemother. 35:590–593.
the corresponding NsiI deletion clones are shown on the right-hand
4. CLSI. 2008. Performance standards for antimicrobial disk and dilution sus-
ceptibility test for bacteria isolated from animals; approved standard—third
APRAMYCIN RESISTANCE GENE apmA IN MRSA ST398
edition. CLSI document M31-A3. Clinical and Laboratory Standards Insti-
Staphylococcus aureus and Staphylococcus pseudintermedius detected in the
BfT-GermVet monitoring programme 2004–2006 in Germany. J. Antimi-
5. Davies, J., and S. O’Connor. 1978. Enzymatic modification of aminoglyco-
crob. Chemother. 61:282–285.
side antibiotics: 3-N-acetyltransferase with broad specificity that determines
19. Schwarz, S., C. Kehrenberg, B. Doublet, and A. Cloeckaert. 2004. Molecular
resistance to the novel aminoglycoside apramycin. Antimicrob. Agents Che-
basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Mi-
mother. 14:69–72.
crobiol. Rev. 28:519–542.
6. de Neeling, A. J., M. J. van den Broek, E. C. Spalburg, M. G. van Santen-
20. Smith, T. C., M. J. Male, A. L. Harper, J. S. Kroeger, G. P. Tinkler, E. D. Verheuvel, W. D. Dam-Deisz, H. C. Boshuizen, A. W. van de Giessen, E. van Moritz, A. W. Capuano, L. A. Herwaldt, and D. J. Diekema. 2009. Methicil- Duijkeren, and X. W. Huijsdens. 2007. High prevalence of methicillin resis-
lin-resistant Staphylococcus aureus (MRSA) strain ST398 is present in mid-
tant Staphylococcus aureus in pigs. Vet. Microbiol. 122:366–372.
western U.S. swine and swine workers. PLoS One 4:e4258.
7. Denis, O., C. Suetens, M. Hallin, B. Catry, I. Ramboer, M. Dispas, G.
21. Threlfall, E. J., B. Rowe, J. L. Ferguson, and L. R. Ward. 1986. Character- Willems, B. Gordts, P. Butaye, and M. J. Struelens. 2009. Methicillin-resis-
ization of plasmids conferring resistance to gentamicin and apramycin in
tant Staphylococcus aureus ST398 in swine farm personnel, Belgium. Emerg.
strains of Salmonella typhimurium phage type 204c isolated in Britain. J. Hyg.
Infect. Dis. 15:1098–1101.
(Lond.) 97:419–426.
8. Feßler, A., C. Scott, K. Kadlec, R. Ehricht, S. Monecke, and S. Schwarz.
22. van de Giessen, A. W., M. G. van Santen-Verheuvel, P. D. Hengeveld, T.
2010. Characterization of methicillin-resistant Staphylococcus aureus ST398
Bosch, E. M. Broens, and C. B. Reusken. 2009. Occurrence of methicillin-
from cases of bovine mastitis. J. Antimicrob. Chemother. 65:619–625.
resistant Staphylococcus aureus in rats living on pig farms. Prev. Vet. Med.
9. Kadlec, K., R. Ehricht, S. Monecke, U. Steinacker, H. Kaspar, J. Mankertz, 91:270–273. and S. Schwarz. 2009. Diversity of antimicrobial resistance pheno- and
23. Vanderhaeghen, W., T. Cerpentier, C. Adriaensen, J. Vicca, K. Hermans,
genotypes of methicillin-resistant Staphylococcus aureus ST398 from dis-
and P. Butaye. 2010. Methicillin-resistant Staphylococcus aureus (MRSA)
eased swine. J. Antimicrob. Chemother. 64:1156–1164.
ST398 associated with clinical and subclinical mastitis in Belgian cows. Vet.
10. Kadlec, K., and S. Schwarz. 2009. Identification of a novel trimethoprim
Microbiol. 144:166–171.
resistance gene, dfrK, in a methicillin-resistant Staphylococcus aureus ST398
24. van Duijkeren, E., M. D. Jansen, S. C. Flemming, H. de Neeling, J. A.
strain and its physical linkage to the tetracycline resistance gene tet(L). Wagenaar, A. H. W. Schoormans, A. van Nes, and A. C. Fluit. 2007. Methi-
Antimicrob. Agents Chemother. 53:776–778.
cillin-resistant Staphylococcus aureus in pigs with exudative epidermitis.
11. Kadlec, K., and S. Schwarz. 2009. Identification of a novel ABC transporter
Emerg. Infect. Dis. 13:1408–1410.
gene, vga(C), located on a multiresistance plasmid from a porcine methicil-
25. van Duijkeren, E., M. Moleman, M. M. Sloet van Oldruitenborgh-Ooster-
lin-resistant Staphylococcus aureus ST398 strain. Antimicrob. Agents Che-
baan, J. Multem, A. Troelstra, A. C. Fluit, W. J. van Wamel, D. J. Houwers,
mother. 53:3589–3591. A. J. de Neeling, and J. A. Wagenaar. 2010. Methicillin-resistant Staphylo-
12. Kadlec, K., and S. Schwarz. 2010. Identification of a plasmid-borne resis- coccus aureus in horses and horse personnel: an investigation of several
tance gene cluster comprising the resistance genes erm(T), dfrK, and tet(L) in
outbreaks. Vet. Microbiol. 141:96–102.
a porcine methicillin-resistant Staphylococcus aureus ST398 strain. Antimi-
26. Voss, A., F. Loeffen, J. Bakker, C. Klaassen, and M. Wulf. 2005. Methicillin-
crob. Agents Chemother. 54:915–918.
resistant Staphylococcus aureus in pig farming. Emerg. Infect. Dis. 11:1965–
13. Kehrenberg, C., C. Cuny, B. Strommenger, S. Schwarz, and W. Witte. 2009.
Methicillin-resistant and -susceptible Staphylococcus aureus strains of clonal
27. Walther, B., S. Monecke, C. Ruscher, A. W. Friedrich, R. Ehricht, P. Slick-
lineages ST398 and ST9 from swine carry the multidrug resistance gene cfr. ers, A. Soba, C. G. Wleklinski, L. H. Wieler, and A. Lu ¨bke-Becker. 2009.
Antimicrob. Agents Chemother. 53:779–781.
Comparative molecular analysis substantiates zoonotic potential of equine
¨thje, P., and S. Schwarz. 2006. Antimicrobial resistance of coagulase-
methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 47:704–710.
negative staphylococci from bovine subclinical mastitis with particular ref-
28. Witte, W., B. Strommenger, C. Stanek, and C. Cuny. 2007. Methicillin-
erence to macrolide-lincosamide resistance phenotypes and genotypes. J.
resistant Staphylococcus aureus ST398 in humans and animals, Central Eu-
Antimicrob. Chemother. 57:966–969.
rope. Emerg. Infect. Dis. 13:255–258.
15. Nemati, M., K. Hermans, U. Lipinska, O. Denis, A. Deplano, M. Struelens,
29. Wray, C., R. W. Hedges, K. P. Shannon, and D. E. Bradley. 1986. Apramycin L. A. Devriese, F. Pasmans, and F. Haesebrouck. 2008. Antimicrobial resis-
and gentamicin resistance in Escherichia coli and salmonellas isolated from
tance of old and recent Staphylococcus aureus isolates from poultry: first
farm animals. J. Hyg. (Lond.) 97:445–456.
detection of livestock-associated methicillin-resistant strain ST398. Antimi-
30. Wulf, M. W., E. Tiemersma, J. Kluytmans, D. Bogaers, A. C. Leenders,
crob. Agents Chemother. 52:3817–3819. M. W. Jansen, J. Berkhout, E. Ruijters, D. Haverkate, M. Isken, and A. Voss.
16. Nienhoff, U., K. Kadlec, I. F. Chaberny, J. Verspohl, G.-F. Gerlach, S.
2008. MRSA carriage in healthcare personnel in contact with farm animals. Schwarz, D. Simon, and I. Nolte. 2009. Transmission of methicillin-resistant
J. Hosp. Infect. 70:186–190. Staphylococcus aureus strains between humans and dogs: two case reports. J.
31. Yates, C. M., M. C. Pearce, M. E. Woolhouse, and S. G. Amyes. 2004. High
Antimicrob. Chemother. 64:660–662.
frequency transfer and horizontal spread of apramycin resistance in calf
17. Pomba, C., F. Baptista, N. Couto, F. Louc ˜o, and H. Hasman. 2010. Methi-
faecal Escherichia coli. J. Antimicrob. Chemother. 54:534–537.
cillin-resistant Staphylococcus aureus CC398 isolates with indistinguishable
32. Zhang, X. Y., L. J. Ding, and M. Z. Fan. 2009. Resistance patterns and
ApaI restriction patterns in colonized and infected pigs and humans. J.
detection of aac(3)-IV gene in apramycin-resistant Escherichia coli isolated
Antimicrob. Chemother. 65:2479–2481.
from farm animals and farm workers in northeastern of China. Res. Vet. Sci.
18. Schwarz, S., K. Kadlec, and B. Strommenger. 2008. Methicillin-resistant 87:449–454.
Cannabinoids 2006;1(1):10-14 [Versión española] Mini-revisión Los cannabinoides y el sistema endocannabinoide Franjo Grotenhermen nova-Institut, Goldenbergstraße 2, D-50354 Hürth, Germany El cuerpo humano posee sitios específicos de acople para los cannabinoides en la superficie de muchos tipos de células y nuestro organismo produce varios endocannabinoides, derivados de áci-dos
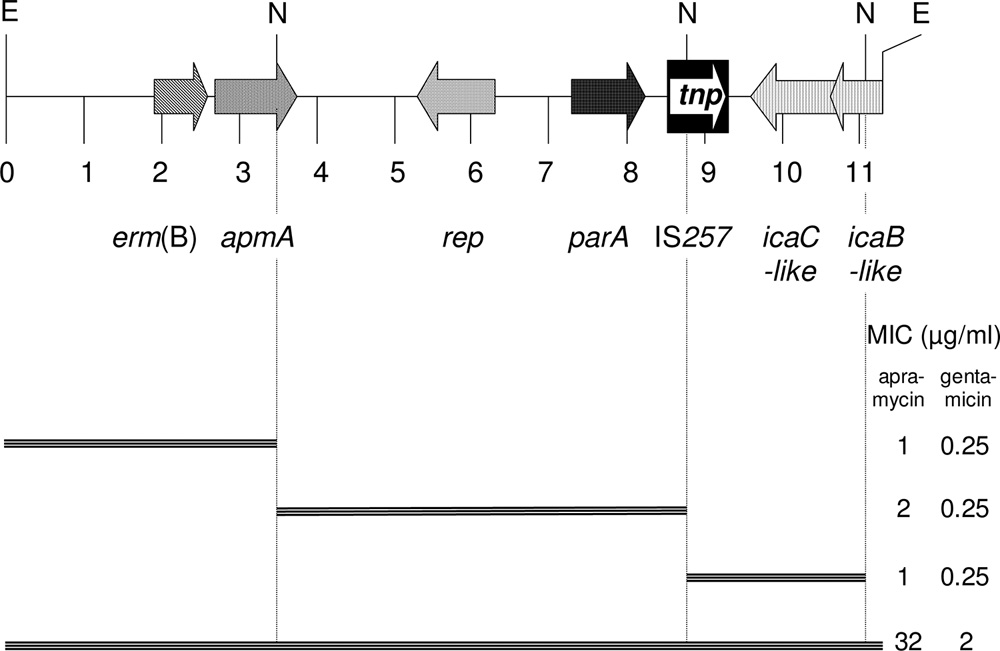 TABLE 1. Comparative analysis of the bovine MRSA ST398 isolate 11, S. aureus RN4220, and the S. aureus RN4220
apmA, erm(B), tet(L), tet(M), tet(K),
dfrK, aadD, mecA, blaZ
apmA, erm(B), tet(L), dfrK, aadD
a APR, apramycin; GEN, gentamicin; ERY, erythromycin; CLI, clindamycin; TET, tetracycline; TMP, trimethoprim; KAN, kanamycin; NEO, neomycin; OXA,
suggested that recombination events via this insertion se-
TTTC-3Ј) (annealing temperature, 52°C; amplicon size, 656
quence have occurred. A complete reading frame for a
bp) was developed and applied to MRSA ST398 isolates.
TABLE 1. Comparative analysis of the bovine MRSA ST398 isolate 11, S. aureus RN4220, and the S. aureus RN4220
apmA, erm(B), tet(L), tet(M), tet(K),
dfrK, aadD, mecA, blaZ
apmA, erm(B), tet(L), dfrK, aadD
a APR, apramycin; GEN, gentamicin; ERY, erythromycin; CLI, clindamycin; TET, tetracycline; TMP, trimethoprim; KAN, kanamycin; NEO, neomycin; OXA,
suggested that recombination events via this insertion se-
TTTC-3Ј) (annealing temperature, 52°C; amplicon size, 656
quence have occurred. A complete reading frame for a
bp) was developed and applied to MRSA ST398 isolates.